
Unit 2: Bacteriology
06
Lecture 2 – The Morphology & Fine
structure of bacteria
The main characteristics that distinguish
prokaryotes from eukaryotes are the
following:
1) Eukaryotic cells are generally more complex than
prokaryotic cells.
2) DNA is enclosed in a nuclear membrane and is associated
with histones and other proteins only in eukaryotes.
3) Organelles are membrane-bound in eukaryotes.
4) Prokaryotes divide by binary fission whereas eukaryotes
divide by mitosis.
5) Some structures are absent in prokaryotes: for example,
Golgi complex, endoplasmic reticulum, mitochondria, and
chloroplasts.
There are more other differences between prokaryotes
and eukaryotes you can find them in reference text books.
The Morphology of Bacteria
Bacteria differ from other single-cell microorganisms in
both their cell structure and size, which varies from 0.3–5
µm. Magnifications of 500–1000X—close to the
resolution limits of light microscopy—are required to
obtain useful images of bacteria. Another problem is that
the structures of objects the size of bacteria offers little
visual contrast. Techniques like phase contrast and dark
field microscopy, both of which allow for live cell
observation, are used to overcome this difficulty.
Chemical-staining techniques are also used, but the
prepared specimens are dead. Bacteria have three basic
forms: cocci, straight rods, and curved or spiral rods.
Cocci are spherical bacteria. Those found in grapelike
clusters as in this picture are staphylococci (Scanning
electron microscopy (SEM)).
The straight rod bacteria with rounded ends
Shown here are coli bacteria (SEM).
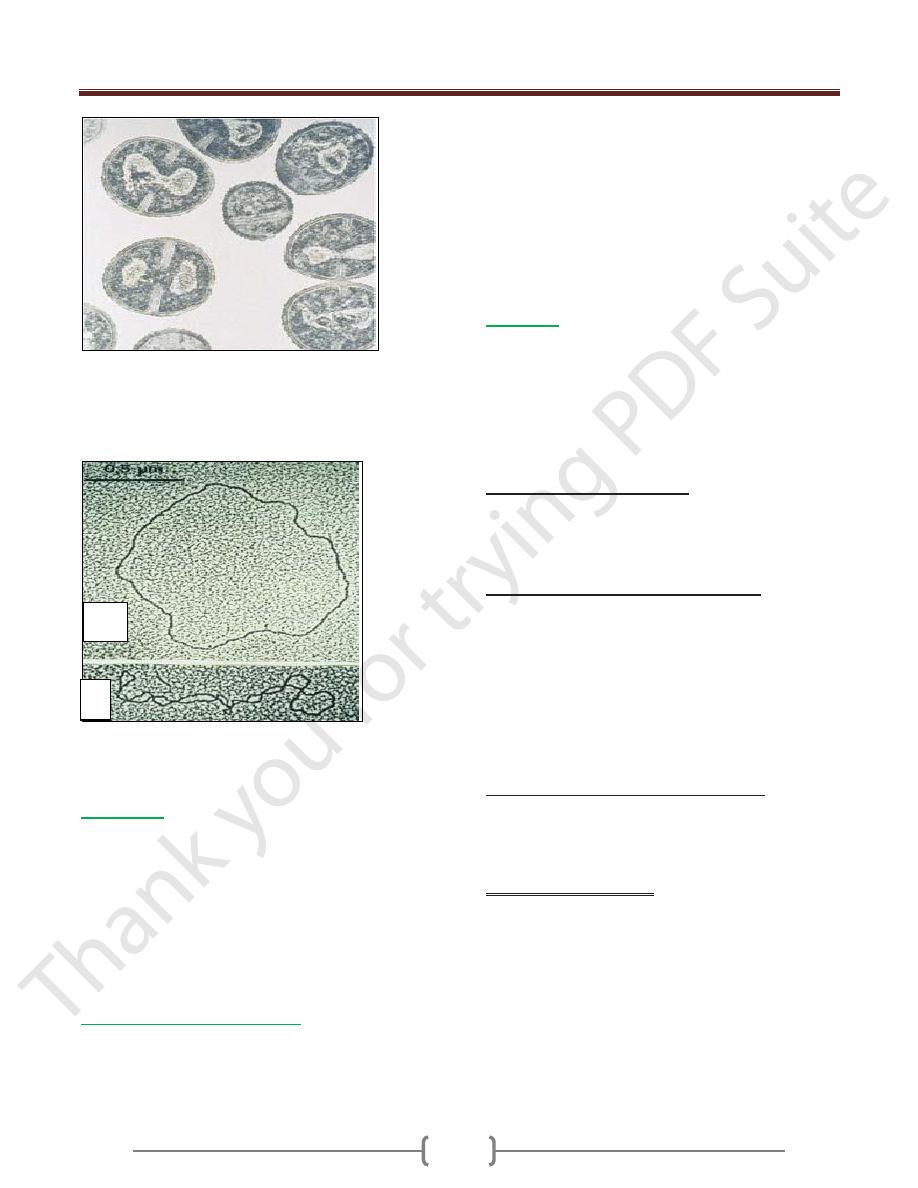
Spirilla, in this case borrelia are spiral
bacteria (light microscopy (LM), Giemsa stain).
The Structures of Bacterial cell
The Nucleoid and plasmids
The prokaryote
Nucleoid ( the equivalent of the
eukaryotic Nucleus)
consists of a very thin, long,
circular DNA molecular double strand that is not
surrounded by a membrane and localized in the
cytoplasm. In E. coli (and probably in all bacteria), it
takes the form of a single circular molecule of DNA. The
genome of E. coli comprises 4.63 X106 base pairs (bp)
that code for 4288 different proteins. The genomic
sequence of many bacteria is known. The plasmids are
nonessential genetic structures. These circular, twisted
DNA molecules are 100–1000X smaller than the nucleoid
genome structure and reproduce autonomously .The
plasmids of human pathogen bacteria often bear important
genes determining the phenotype of their cells (resistance
genes, virulence genes).
Unit 2: Bacteriology
06
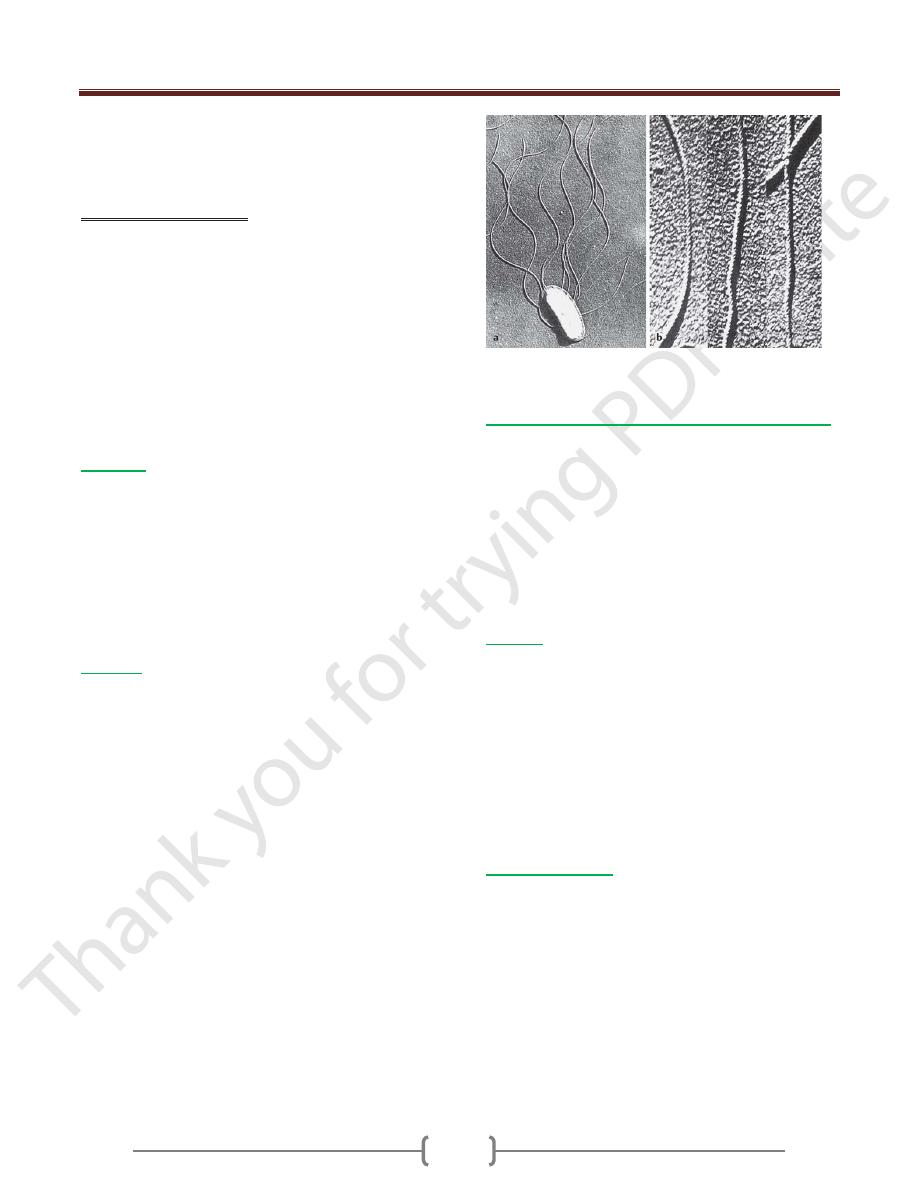
The nucleoid (nucleus equivalent) of bacteria consists of a
tangled circular DNA molecule without a nuclear
membrane.
Transmission electron microscopy (TEM) image of
staphylococci
A) Open circular form (OC). The result of a rupture in
one of the two nucleic acid strands. B) Twisted (CCC =
covalently closed circular), native form (TEM image).
Cytoplasm
The cytoplasm contains a large number of solute (low-
and high-molecularweight) substances, RNA and
ribosomes (approximately 20 000 per cell). Bacteria have
70S ribosomes comprising 30S and 50S subunits.
Bacterial ribosomes function as the organelles for protein
synthesis. The cytoplasm is also frequently used to store
reserve substances (glycogen depots, polymerized
metaphosphates, lipids).
The Cytoplasmic Membrane
This elementary membrane, also known as the plasma
membrane, is
a 40–80 A ˚-thick semipermeable
membrane
. It is basically a double layer of phospholipids
with numerous proteins integrated into its structure. The
most important of these membrane proteins are
permeases, enzymes for the biosynthesis of the cellwall,
transfer proteins for secretion of extracellular proteins,
sensor or signal proteins, and respiratory chain enzymes.
In electron microscopic images of Gram-positive bacteria,
the mesosomes appear as structures bound to the
membrane. How they function and what role they play
remain to be clarified.
Cell Wall
The tasks of the complex bacterial cellwall are to protect
the protoplasts from external noxae, to withstand and
maintain the osmotic pressure gradient between the cell
interior and the extracellular environment (with internal
pressures as high as 500–2000 kPa), to give the cell its
outer form and to facilitate communication with its
surroundings.
Peptidoglycan (syn. murein).
The most important
structural element of the wall is peptidoglycan, a netlike
polymer material surrounding the entire cell (sacculus). It
is made up of polysaccharide chains crosslinked by
peptides.
The cell wall of Gram-positive bacteria
. The
murein sacculus may consist of as many as 40 layers (15–
80 nm thick) and account for as much as 30% of the dry
mass of the cell wall. The membrane lipoteichoic acids
are anchored in the cytoplasmic membrane, whereas the
cell wall teichoic acids are covalently coupled to the
murein. The physiological role of the teichoic acids is not
known in detail; possibly they regulate the activity of the
autolysins that steer growth and transverse fission
processes in the cell.
The cell wall of Gram-negative bacteria
. Here, the
murein is only about 2 nm thick and contributes up to 10%
of the dry cell wall mass. The outer membrane is the salient
structural element. It contains numerous proteins (50% by
mass) as well as the medically critical lipopolysaccharide.
Outer membrane proteins
—
OmpA (outer membrane protein A) and the murein
lipoprotein form a
bond between outer membrane and murein.
—
Porins, proteins that form pores in the outer
membrane, allow passage of hydrophilic, low-molecular-
weight substances into the periplasmic space.
—
Outer membrane-associated proteins constitute
specific structures that enable bacteria to attach to host
cell receptors.
A
B
Unit 2: Bacteriology
06
—
A number of Omps are transport proteins.
Examples include the LamB proteins for maltose transport
and FepA for transport of the siderophore ferric (Fe3+)
enterochelin in E. coli .
Lipopolysaccharide (LPS).
This molecular complex, also known as endotoxin, is
comprised of the lipoid A, the core polysaccharide, and
the O-specific polysaccharide chain.
—
Lipoid A is responsible for the toxic effect.
—
The O-specific polysaccharide chain is the so-called
O antigen.
L-forms (L = Lister Institute). The L-forms are bacteria
with murein defects, e.g., resulting from the effects of
betalactam antibiotics. L-forms are highly unstable when
subjected to osmotic influences.
Capsule
Many pathogenic bacteria make use of extracellular
enzymes to synthesize a polymer that forms a layer
around the cell: the capsule. The capsule protects bacterial
cells from phagocytosis. The capsule of most bacteria
consists of a polysaccharide. The bacteria of a single
species can be classified in different capsular serovars (or
serotypes) based on the fine chemical structure of this
polysaccharide.
Flagella
Flagella give bacteria the ability to move about actively.
The flagella (singular flagellum) are made up of a class of
linear proteins called flagellins. Flagellated bacteria
display various flagellar arrangements ranging from
monotrichous (polar flagellum; e.g., Vibrio coma),
lophotrichous (bundle of flagella at one end of the cell;
e.g., Spirillum volutans), to peritrichous (several flagella
distributed around the cell; e.g., Escherichia coli)
.
Together with the O antigens, they are used to classify
bacteria in serovars.
a
Flagellated bacterial cell (SEM, 13 000!). b Helical
structure of bacterial flagella (SEM, 77 000X).
Attachment Pili (Fimbriae), Conjugation Pili
Many Gram-negative bacteria possess thin microfibrils
made of proteins (0.1–1.5 nm thick, 4–8 nm long), the
attachment pili. They are anchored in the outer membrane
of the cell wall and extend radially from the surface.
Using these structures, bacteria are capable of specific
attachment to host cell receptors (ligand—receptor, key—
keyhole). The conjugation pili (syn. sex pili) in Gram-
negative bacteria are required for the process of
conjugation and thus for transfer of conjugative plasmids.
Biofilm
A bacterial biofilm is a structured community of bacterial
cells embedded in a self-produced polymer matrix and
attached to either an inert surface or living tissue. Such
films can develop considerable thickness (mm) . Foreign
body infections are caused by bacteria that form a biofilm
on inert surfaces. The bacteria located deep within such a
biofilm structure are effectively isolated from immune
system cells, antibodies, and antibiotics. The polymers
they secrete are frequently glycosides, from which the
term glycocalyx (glycoside cup) for the matrix is derived.
Bacterial Spores
Bacterial spores (endospores) are purely dormant life
forms. Their development from bacterial cells in a
“vegetative” state does not involve assimilation of
additional external nutrients. They are spherical to oval in
shape and are characterized by a thick spore wall & a high
level of resistance to chemical & physical noxae. Among
human pathogen bacteria, only the genera Clostridium
and Bacillus produce spores. The heat resistance of these
spores is their most important quality from a medical
point of view, since heat sterilization procedures require
very high temperatures to kill them effectively.