
fact that we experience these different modalities of sensation, nerve fibers
of sensation. Yet despite the
sight, sound, and so forth—is called a
Each of the principal types of sensation that we can experience—pain, touch,
enough to damage the tissues.
of the body fluids but have never been known to respond to sound. Finally, pain
on the eyeballs, or chemical changes in the blood. The osmoreceptors of the
are almost completely nonresponsive to normal ranges of heat, cold, pressure
stimuli. Thus, the rods and cones of the eyes are highly responsive to light but
sitivities.” That is, each type of receptor is highly sensitive to one type of stimulus
tors detect different types of sensory stimuli? The answer is, by “differential sen-
The first question that must be answered is, how do two types of sensory recep-
Differential Sensitivity of Receptors
types of mechanoreceptors found in the skin or in deep tissues of the body.
tion to the sensory systems that they subserve. Figure 46–1 shows some of the
which receptors operate. Other receptors are discussed in other chapters in rela-
primarily peripheral mechanoreceptors, to illustrate some of the principles by
In this chapter, we discuss the function of a few specific types of receptors,
up the chemistry of the body.
body fluids, carbon dioxide concentration, and perhaps other factors that make
mouth, smell in the nose, oxygen level in the arterial blood, osmolality of the
, which detect taste in the
chemoreceptors
on the retina of the eye; and (5)
, which detect light
cal damage or chemical damage; (4)
(pain receptors), which detect damage occurring in the tissues, whether physi-
temperature, some receptors detecting cold and others warmth; (3)
, which detect changes in
tissues adjacent to the receptor; (2)
, which detect mechanical compression or stretching of the receptor or of
mechanorecep-
shows that there are five basic types of sensory receptors: (1)
Table 46–1 lists and classifies most of the body’s sensory receptors. This table
Types of Sensory Receptors and the Sensory
sound, light, pain, cold, and warmth. The purpose of
Sensory Receptors, Neuronal
C
H
A
P
T
E
R
4
6
572
Circuits for Processing Information
Input to the nervous system is provided by sensory
receptors that detect such sensory stimuli as touch,
this chapter is to discuss the basic mechanisms by
which these receptors change sensory stimuli into
nerve signals that are then conveyed to and
processed in the central nervous system.
Stimuli They Detect
tors
thermoreceptors
nociceptors
electromagnetic receptors
for which it is designed and yet is almost nonresponsive to other types of sensory
supraoptic nuclei in the hypothalamus detect minute changes in the osmolality
receptors in the skin are almost never stimulated by usual touch or pressure
stimuli but do become highly active the moment tactile stimuli become severe
Modality of Sensation—The “Labeled Line” Principle
modality

tor, its immediate effect is to change the membrane
Whatever the type of stimulus that excites the recep-
Stimuli into Nerve Impulses
Transduction of Sensory
This specificity of nerve fibers for transmitting only
areas.
terminate in the vision areas of the brain, fibers from
the brain. Similarly, fibers from the retina of the eye
or in any other way, the person perceives touch
the person perceives pain. Likewise, if a touch fiber is
by damage to the tissue cells. In all these instances,
of the fiber, or stimulation of the pain nerve ending
can be electricity, overheating of the fiber, crushing
of what type of stimulus excites the fiber. The stimulus
is stimulated, the person perceives pain regardless
to which the fiber leads. For instance, if a pain fiber
specific point in the central nervous system, and
The answer is that each nerve tract terminates at a
transmit only impulses. Therefore, how is it that
Sensory Receptors, Neuronal Circuits for Processing Information
Chapter 46
573
different nerve fibers transmit different modalities of
sensation?
the type of sensation felt when a nerve fiber is stimu-
lated is determined by the point in the nervous system
stimulated by electrical excitation of a touch receptor
because touch fibers lead to specific touch areas in
the ear terminate in the auditory areas of the brain,
and temperature fibers terminate in the temperature
one modality of sensation is called the labeled line
principle.
Local Electrical Currents at Nerve
Endings—Receptor Potentials
All sensory receptors have one feature in common.
Free nerve
endings
Expanded tip
receptor
Tactile hair
Pacinian
corpuscle
Meissner’s
corpuscle
Krause’s
corpuscle
Ruffini’s
end-organ
Golgi tendon
apparatus
Muscle
spindle
Several types of somatic sensory nerve endings.
Figure 46–1
Table 46–1
Blood glucose, amino acids, fatty acids
Taste
V. Chemoreceptors
Vision
IV. Electromagnetic receptors
Free nerve endings
III. Nociceptors
Warm receptors
Warmth
II. Thermoreceptors
Vestibular receptors
Ruffini’s endings
Free nerve endings
Krause’s corpuscles
Meissner’s corpuscles
Ruffini’s endings
Merkel’s discs
Free nerve endings
I. Mechanoreceptors
Classification of Sensory Receptors
Skin tactile sensibilities (epidermis and dermis)
Expanded tip endings
Plus several other variants
Spray endings
Encapsulated endings
Hair end-organs
Deep tissue sensibilities
Expanded tip endings
Spray endings
Encapsulated endings
Pacinian corpuscles
Plus a few other variants
Muscle endings
Muscle spindles
Golgi tendon receptors
Hearing
Sound receptors of cochlea
Equilibrium
Arterial pressure
Baroreceptors of carotid sinuses and aorta
Cold
Cold receptors
Pain
Rods
Cones
Receptors of taste buds
Smell
Receptors of olfactory epithelium
Arterial oxygen
Receptors of aortic and carotid bodies
Osmolality
Neurons in or near supraoptic nuclei
Blood CO
2
Receptors in or on surface of medulla and in aortic and
carotid bodies
Receptors in hypothalamus
of a pacinian corpuscle. Note that the amplitude
mechanical compression (increasing “stimulus
Figure 46–4 shows the changing amplitude of the
izes the fiber membrane at this node, which then sets
the pacinian corpuscle, the local current flow depolar-
node of Ranvier, which itself lies inside the capsule of
arrows, that spreads along the nerve fiber. At the first
of current flow, shown by the
“receptor potential.” The receptor potential in turn
increased positivity inside the fiber, which is the
diffuse to the interior of the fiber. This creates
brane, allowing positively charged sodium ions to
has been deformed by compression of the corpuscle,
cle. Observe the small area of the terminal fiber that
The figure also shows the mechanism by which a
puscle to enter a peripheral sensory nerve.
central fiber inside the capsule is unmyelinated, but
layers but one have been removed. The tip of the
Now study Figure 46–3, which shows only the
indent, or otherwise deform the central fiber.
anywhere on the outside of the corpuscle will elongate,
tiple concentric capsule layers, so that compression
extending through its core. Surrounding this are mul-
46–1. Note that the corpuscle has a central nerve fiber
cal structure of the pacinian corpuscle shown in Figure
The student should at this point restudy the anatomi-
receptor potential rises above the threshold level, the
trated in Figure 46–2. Note also that the more the
to the receptor, then action potentials occur, as illus-
When
becomes maximally permeable to sodium ions.
high intensity of sensory stimulus. This is about the
100 millivolts, but this level occurs only at an extremely
The maximum
change in membrane permeability of the receptor,
types of known sensory receptors. In all instances, the
and allows ions to flow through membrane channels.
retinal visual receptor, which either directly or indi-
effects of electromagnetic radiation, such as light on a
alters the permeability of the membrane; or (4) by the
change of the temperature of the membrane, which
the membrane, which also opens ion channels; (3) by
opens ion channels; (2) by application of a chemical to
receptor, which stretches the receptor membrane and
tor potentials: (1) by mechanical deformation of the
electrical potential of the receptor. This change in
The Nervous System: A. General Principles and Sensory Physiology
574
Unit IX
potential is called a receptor potential.
Mechanisms of Receptor Potentials.
Different receptors
can be excited in one of several ways to cause recep-
rectly changes the receptor membrane characteristics
It will be recognized that these four means of exciting
receptors correspond in general with the different
basic cause of the change in membrane potential is a
which allows ions to diffuse more or less readily
through the membrane and thereby to change the
transmembrane potential.
Maximum Receptor Potential Amplitude.
amplitude of most sensory receptor potentials is about
same maximum voltage recorded in action potentials
and is also the change in voltage when the membrane
Relation of the Receptor Potential to Action Potentials.
the receptor potential rises above the threshold for
eliciting action potentials in the nerve fiber attached
greater becomes the action potential frequency.
Receptor Potential of the Pacinian Corpuscle—
An Example of Receptor Function
central fiber of the pacinian corpuscle after all capsule
the fiber does become myelinated (the blue sheath
shown in the figure) shortly before leaving the cor-
receptor potential is produced in the pacinian corpus-
and note that ion channels have opened in the mem-
induces a local circuit
off typical action potentials that are transmitted along
the nerve fiber toward the central nervous system.
Relation Between Stimulus Intensity and the Receptor Poten-
tial.
receptor potential caused by progressively stronger
strength”) applied experimentally to the central core
60
80
100 120 140
0
10
20
30
40
Membrane potential (millivolts)
Milliseconds
Resting membrane potential
Receptor potential
Threshold
Action potentials
+
30
-
30
-
60
-
90
0
when the receptor potential rises above threshold level.
Typical relation between receptor potential and action potentials
Figure 46–2
- - - -
- - - -
+ + + + + + + + + + + + + + +
+ + + + + + + + + + + + + + +
+
+
+
+
+
+
+
+
+
+ + + +
+
+ + + +
+
+
+
+
+
+
+
+
+
+
+
+
+
+ + + +
+ + + +
Deformed
area
Receptor potential
Action
potential
Node of
Ranvier
Excitation and inactivation in a receptor membrane. Ann N Y Acad
duced in a pacinian corpuscle. (Modified from Loëwenstein WR:
Excitation of a sensory nerve fiber by a receptor potential pro-
Figure 46–3
Sci 94:510, 1961.)
tial is no longer elicited. Thus, the receptor potential
the corpuscle redistributes, so that the receptor poten-
within a few hundredths of a second, the fluid within
fiber, thus eliciting a receptor potential. However,
applied to one side of the corpuscle, this force is
ways. First, the pacinian corpuscle is a viscoelastic
corpuscle. Adaptation occurs in this receptor in two
In the case of the mechanoreceptors, the receptor
instance, in the eye, the rods and cones adapt by chang-
receptor potential is an individual property. For
tor, in much the same way that development of a
The mechanism of
completely.
pain receptors, for instance—probably never adapt
tid and aortic baroreceptors. Conversely, some of
2 days, which is the adaptation time for many caro-
adapting” receptors. The longest measured time for
or days to do so, for which reason they are called “non-
ally adapt almost completely, but some require hours
mechanoreceptors
the hairs adapt to extinction within a second or more.
dredths of a second, and the receptors at the bases of
corpuscles adapt to “extinction” within a few hun-
greater extent than others. For example, the pacinian
Furthermore, some sensory receptors adapt to a far
muscle spindle receptors adapt slowly.
within a second or so, whereas some joint capsule and
types of receptors. Note that the pacinian corpuscle
Figure 46–5 shows typical adaptation of certain
continuous sensory stimulus is applied, the receptor
stant stimulus after a period of time. That is, when a
Adaptation of Receptors
weak to very intense.
tor to have an extreme range of response, from very
sensory experience is extreme. This allows the recep-
cable to almost all sensory receptors. It allows the
This is an exceedingly important principle that is appli-
additional increase in numbers of action potentials.
in Figure 46–4, one can see that very intense stimula-
potential. Putting this principle together with the data
In turn, the
Sensory Receptors, Neuronal Circuits for Processing Information
Chapter 46
575
increases rapidly at first but then progressively less
rapidly at high stimulus strength.
frequency of repetitive action potentials
transmitted from sensory receptors increases approx-
imately in proportion to the increase in receptor
tion of the receptor causes progressively less and less
receptor to be sensitive to very weak sensory experi-
ence and yet not reach a maximum firing rate until the
Another characteristic of all sensory receptors is that
they adapt either partially or completely to any con-
responds at a high impulse rate at first and then at a
progressively slower rate until finally the rate of action
potentials decreases to very few or often to none at all.
adapts extremely rapidly and hair receptors adapt
It is probable that all other
eventu-
complete adaptation of a mechanoreceptor is about
the nonmechanoreceptors—the chemoreceptors and
Mechanisms by Which Receptors Adapt.
receptor adaptation is different for each type of recep-
ing the concentrations of their light-sensitive chemi-
cals (which is discussed in Chapter 50).
that has been studied in greatest detail is the pacinian
structure so that when a distorting force is suddenly
instantly transmitted by the viscous component of the
corpuscle directly to the same side of the central nerve
0
20
40
60
80
100
Amplitude of observed
receptor potential (per cent)
Stimulus strength
(per cent)
0
10
20
30
40
50
60
70
80
90
100
Loëwenstein WR: Excitation and inactivation in a receptor mem-
mechanical stimulus applied to a pacinian corpuscle. (Data from
Relation of amplitude of receptor potential to strength of a
Figure 46–4
brane. Ann N Y Acad Sci 94:510, 1961.)
0
1
2
3
4
5
6
7
8
Impulses per second
Seconds
0
250
Joint capsule receptors
200
150
100
50
Muscle spindle
Hair receptor
Pacinian corpuscle
tion of some receptors and slow adaptation of others.
Adaptation of different types of receptors, showing rapid adapta-
Figure 46–5

fibers into two subgroups; yet
separate the type A
Alternative Classification Used by Sensory Physiologists.
slowly as 0.5 m/sec, requiring about 2 seconds to go from
Conversely, the smallest fibers transmit impulses as
mit impulses at velocities as great as 120 m/sec, a
46–6. Note that a few large myelinated fibers can trans-
the different nerve fiber types are also given in Figure
The sizes, velocities of conduction, and functions of
well as all the postganglionic autonomic fibers.
at low velocities. The C fibers constitute more than one
fibers of spinal nerves. Type C fibers are the
Type A fibers are the typical large and medium-sized
fibers.
, and
and C, and the type A fibers are further subdivided into
general classification, the fibers are divided into types A
fication” of the different types of nerve fibers. In the
is a “general classification” and a “sensory nerve classi-
Shown in Figure 46–6
General Classification of Nerve Fibers.
conducting velocities is between 0.5 and 120 m/sec.
ter, the greater the conducting velocity. The range of
Figure 46–6, nerve fibers come in all sizes between 0.5
that slowly conducting fibers will suffice. As shown in
aching pain, do not need to be transmitted rapidly, so
sensory information, such as that depicting prolonged,
during running. At the other extreme, some types of
the information would be useless. An example of this is
central nervous system extremely rapidly; otherwise,
Different Types of Signals,
Nerve Fibers That Transmit
tion so that the person will not fall. Loss of this pre-
tion of the next second. Therefore, appropriate motor
instance, when one is running, information from the
of movement of the different parts of the body. For
to keep from losing balance. Likewise, recep-
around a curve. Using this information, a person can
later. For instance, the receptors of the semicircular
status is taking place, one can predict in one’s mind the
body.
of rapid tissue deformations, but it is useless for trans-
released. In other words, the pacinian corpuscle is
later, it transmits a signal again when the pressure is
tation is over even though the pressure continues. But
this receptor for a few milliseconds, and then its exci-
puscle, sudden pressure applied to the tissue excites
receptors. Thus, in the case of the pacinian cor-
receptors, or
movement
receptors,
. Therefore, these recep-
change is actually taking place
strength changes. Yet they react strongly
receptors.
to transmit information for many hours, they are called
chemoreceptors of the carotid and aortic bodies.
receptors, (3) baroreceptors of the arterial tree, and (4)
tors of the macula in the vestibular apparatus, (2) pain
its surroundings. For instance, impulses from the
or hours). Therefore, they keep the brain constantly
Strength—The “Tonic” Receptors.
readjustments in the structure of the receptor itself,
receptors. That is, part of the adaptation results from
Presumably, these same two general mechanisms of
or most cell membrane sodium channels, as was
ally to close, an effect that seems to occur for all
the nerve fiber membrane, which means that sodium
progressive “inactivation” of the sodium channels in
dated” to the stimulus. This probably results from
the nerve fiber itself gradually becomes “accommo-
core fiber should continue to be distorted, the tip of
nerve fiber itself. That is, even if by chance the central
, which occurs in the
ian corpuscle, but a much slower one, results from a
The second mechanism of adaptation of the pacin-
compression continues.
The Nervous System: A. General Principles and Sensory Physiology
576
Unit IX
appears at the onset of compression but disappears
within a small fraction of a second even though the
process called accommodation
current flow through the channels causes them gradu-
explained in Chapter 5.
adaptation apply also to the other types of mechano-
and part from an electrical type of accommodation in
the terminal nerve fibril.
Slowly Adapting Receptors Detect Continuous Stimulus
Slowly adapting recep-
tors continue to transmit impulses to the brain as long
as the stimulus is present (or at least for many minutes
apprised of the status of the body and its relation to
muscle spindles and Golgi tendon apparatuses allow
the nervous system to know the status of muscle con-
traction and load on the muscle tendon at each instant.
Other slowly adapting receptors include (1) recep-
Because the slowly adapting receptors can continue
tonic
Rapidly Adapting Receptors Detect Change in Stimulus
Strength—The “Rate Receptors,” “Movement Receptors,”
or “Phasic Receptors.”
Receptors that adapt rapidly
cannot be used to transmit a continuous signal because
these receptors are stimulated only when the stimulus
while a
tors are called rate
phasic
exceedingly important in apprising the nervous system
mitting information about constant conditions in the
Importance of the Rate Receptors—Their Predictive Function.
If one knows the rate at which some change in bodily
state of the body a few seconds or even a few minutes
canals in the vestibular apparatus of the ear detect the
rate at which the head begins to turn when one runs
predict how much he or she will turn within the next
2 seconds and can adjust the motion of the legs ahead
of time
tors located in or near the joints help detect the rates
joint rate receptors allows the nervous system to
predict where the feet will be during any precise frac-
signals can be transmitted to the muscles of the legs to
make any necessary anticipatory corrections in posi-
dictive function makes it impossible for the person to
run.
and Their Physiologic
Classification
Some signals need to be transmitted to or from the
the sensory signals that apprise the brain of the momen-
tary positions of the legs at each fraction of a second
and 20 micrometers in diameter—the larger the diame-
a, b, g
d
myelinated
small unmyelinated nerve fibers that conduct impulses
half of the sensory fibers in most peripheral nerves as
distance in 1 second that is longer than a football field.
the big toe to the spinal cord.
Certain recording techniques have made it possible to
a

intensity of pain. The different gradations of intensity
must be conveyed is signal intensity—for instance, the
Temporal Summation
Nerve Tracts—Spatial and
of Different Intensity in
Transmission of Signals
diameter; they are type C fibers in the general classifi-
Unmyelinated fibers carrying pain, itch, temperature,
-type A fibers in the general classification).
ter; they are
Fibers carrying temperature, crude touch, and pricking
-type A fibers in the general classification).
dles (average about 8 micrometers in diameter; these
and from the flower-spray endings of the muscle spin-
Fibers from most discrete cutaneous tactile receptors
micrometers in diameter; these also are
Fibers from the Golgi tendon organs (average about 16
(average about 17 microns in diameter; these are
Fibers from the annulospiral endings of muscle spindles
fibers. Therefore, the fol-
easily between A
Sensory Receptors, Neuronal Circuits for Processing Information
Chapter 46
577
these same recording techniques cannot distinguish
b and Ag
lowing classification is frequently used by sensory
physiologists:
Group Ia
a-type
A fibers in the general classification).
Group Ib
a-type A
fibers).
Group II
are
b- and g
Group III
pain sensations (average about 3 micrometers in diame-
d
Group IV
and crude touch sensations (0.5 to 2 micrometers in
cation).
One of the characteristics of each signal that always
15
1
10
5
2.0
0.5
6 2.0
0.5
I
II
III
IV
10
5
1 2.0
0.5
Myelinated
Unmyelinated
20
15
Nerve fiber diameter (micrometers)
Conduction velocity (m/sec.)
30
60
80
IB
IA
C
d
g
b
A
a
Muscle spindle
(primary ending)
Muscle spindle
(secondary ending)
Muscle tendon
(Golgi tendon organ)
Hair receptors
Sensory functions
Sensory nerve classification
General classification
Diameter (micrometers)
Skeletal muscle
(type A
a
)
Muscle spindle
(type A
g
)
Sympathetic
(type C)
Motor function
Deep pressure
and touch
Pricking pain
Cold
Warmth
Aching pain
Tickle
Crude touch
and pressure
Vibration
(pacinian corpuscle)
High discrimination touch
(Meissner's expanded tips)
120
20
Figure 46–6
Physiologic classifications and functions of nerve
fibers.
that causes it to process signals in its own unique way,
long pool of neurons.
cephalon, pons, and medulla. Also, the entire dorsal
specific nuclei in the thalamus, cerebellum, mesen-
ered to be a single large neuronal pool. Other neuronal
instance, the entire cerebral cortex could be consid-
few neurons, while others have vast numbers. For
to millions of neuronal pools; some of these contain
The central nervous system is composed of thousands
of Signals in Neuronal Pools
Transmission and Processing
by the nerve fiber.
. Figure 46–8 demonstrates this,
of nerve impulses in each fiber, which is called
Temporal Summation.
stronger signals spread to more and more fibers. This
gressively more fibers being stimulated. Thus, the
moderate stimulus and a strong stimulus, with pro-
stimulated weakly (half-red fibers). The other two
red-colored fiber), whereas several adjacent fibers are
stimulus, with only a single nerve fiber in the middle
from the skin area. To the left is the effect of a weak
Thus, the lower part of Figure 46–7 shows three
field is much greater than at the periphery.
than when it is in the periphery of the field, because
center of the receptive field of a particular pain fiber,
fibers simultaneously. When the pinprick is in the
from other pain fibers. Therefore, a pinprick of the skin
diminishes toward the periphery. One can also see
of that fiber. The
This area is called the
an area of skin as large as 5 centimeters in diameter.
that serve as pain receptors. The entire
innervated by a large number of parallel pain fibers.
numbers of fibers. This figure shows a section of skin
, whereby increasing signal
Figure 46–7 shows the phenomenon
respectively,
spatial summation and temporal
along a single fiber. These two mechanisms are called,
The Nervous System: A. General Principles and Sensory Physiology
578
Unit IX
can be transmitted either by using increasing numbers
of parallel fibers or by sending more action potentials
summation.
Spatial Summation.
of spatial summation
strength is transmitted by using progressively greater
Each of these arborizes into hundreds of minute free
nerve endings
cluster of fibers from one pain fiber frequently covers
receptor field
number of endings is large in the center of the field but
from the figure that the arborizing fibrils overlap those
usually stimulates endings from many different pain
the degree of stimulation of that fiber is far greater
the number of free nerve endings in the middle of the
views of the cross section of the nerve bundle leading
of the bundle stimulated strongly (represented by the
views of the nerve cross section show the effect of a
is the phenomenon of spatial summation.
A second means for transmitting
signals of increasing strength is by increasing the fre-
quency
temporal summation
showing in the upper part a changing strength of signal
and in the lower part the actual impulses transmitted
pools include the different basal ganglia and the
gray matter of the spinal cord could be considered one
Each neuronal pool has its own special organization
Weak
stimulus
Moderate
stimulus
Skin
Nerve
Pin
Strong
stimulus
Pattern of stimulation of pain fibers in a nerve leading from an area
Figure 46–7
of skin pricked by a pin. This is an example of spatial summation.
Strength of signal
(impulses per second)
Impulses
Time
0
80
60
40
20
of nerve impulses, showing the strength of signal (
Translation of signal strength into a frequency-modulated series
Figure 46–8
above) and the
separate nerve impulses (below). This is an example of temporal
summation.

toward its edges.
endings in the center; it becomes progressively less
The degree of inhibition in the center
inhibitory zone.
exciting them. This is the opposite of facilitation, and
that some incoming fibers inhibit neurons, rather than
We must also remember
subliminal zone
zone
facilitated zone,
are facilitated but not excited, and these areas are
. To each side, the neurons
liminal zone
excited zone
of the incoming fiber, also called the
charge zone
incoming fiber. Therefore, this is said to be the
circled area, all the neurons are stimulated by the
portion of the field in this figure, designated by the
bution “field,” as shown in Figure 46–10. In the central
Figure 46–9 represents a highly condensed version
are subthreshold, but facilitating, stimuli.
is a suprathreshold stimulus, and the stimuli to
, the stimulus to
Similarly, for
through other incoming nerve fibers. Therefore, the
theless, discharge of these terminals makes both these
b and c, but not enough to cause excitation. Never-
; it is also called a
from input fiber 1 to this neuron is said to be an
to discharge. The stimulus
input fiber 1
on each one of the neurons from each input fiber, he
neurons. If the student counts the number of terminals
46–9, let us assume that six terminals must discharge
cession to cause excitation. For instance, in Figure
action potential in a postsynaptic neuron. Instead,
45, it will be recalled that discharge of a single
From the discussion of synaptic function in Chapter
farther away.
input fiber lie on the nearest neuron in its “field,” but
. Note in Figure
The neuronal area stimulated by each incoming
in the pool. The dendrites usually also arborize and
to thousands of times, providing a thousand or more
fibers to the right. Each input fiber divides hundreds
pool, showing “input” fibers to the left and “output”
Figure 46–9 is
Relaying of Signals Through
the following pages.
have many similar principles of function, described in
despite their differences in function, the pools also
the multitude of functions of the nervous system. Yet
Sensory Receptors, Neuronal Circuits for Processing Information
Chapter 46
579
thus allowing the total consortium of pools to achieve
Neuronal Pools
Organization of Neurons for Relaying Signals.
a schematic diagram of several neurons in a neuronal
terminal fibrils that spread into a large area in the pool
to synapse with dendrites or cell bodies of the neurons
spread hundreds to thousands of micrometers in the
pool.
nerve fiber is called its stimulatory field
46–9 that large numbers of the terminals from each
progressively fewer terminals lie on the neurons
Threshold and Subthreshold Stimuli—Excitation or Facilitation.
excitatory presynaptic terminal almost never causes an
large numbers of input terminals must discharge on
the same neuron either simultaneously or in rapid suc-
almost simultaneously to excite any one of the
or she will see that
has more than enough
terminals to cause neuron a
excitatory stimulus
suprathreshold
stimulus because it is above the threshold required for
excitation.
Input fiber 1 also contributes terminals to neurons
neurons more likely to be excited by signals arriving
stimuli to these neurons are said to be subthreshold,
and the neurons are said to be facilitated.
input fiber 2
neuron d
neurons
b and c
of a neuronal pool because each input nerve fiber
usually provides massive numbers of branching termi-
nals to hundreds or thousands of neurons in its distri-
dis-
or
called the
also called the subthreshold
or
.
Inhibition of a Neuronal Pool.
the entire field of the inhibitory branches is called the
of this zone is great because of large numbers of
1
2
d
c
b
a
Basic organization of a neuronal pool.
Figure 46–9
Facilitated zone
Input nerve
fiber
Discharge zone
Facilitated zone
“Discharge” and “facilitated” zones of a neuronal pool.
Figure 46–10

second output pathway from the pool. This type of
(neuron 2), which secretes
excitatory output pathway, but it stimulates an inter-
tion is achieved. The input fiber directly excites the
Figure 46–13 shows the means by which the inhibi-
ling all antagonistic pairs of muscles, and it is called the
ment. This type of circuit is characteristic for control-
spinal cord to cause forward movement of a leg, an
elsewhere. For instance, at the same time that an exci-
Inhibitory Output Signals
Neuronal Circuit with Both Excitatory and
mates, and sorts different types of information.
which the central nervous system correlates, sum-
tion. Convergence is one of the important means by
from different sources, and the resulting response is a
ing from the brain into the spinal cord.Then the signals
cortex, and (4) several other long pathways descend-
another, (3) corticospinal fibers from the cerebral
eral nerve fibers entering the cord, (2) propriospinal
For instance, the interneurons of the
in Figure 46–12
, as shown
to the threshold required for discharge.
tial from a single input terminal. But action potentials
the same neuron. The importance of this is that
. That is, multiple ter-
uniting to excite a single neuron. Figure 46–12
Convergence of Signals
mus, almost all sensory information is relayed both
thalamus and cerebral cortex. Likewise, in the thala-
the lower part of the brain: (1) into the cerebellum and
pool. For instance, information transmitted up the
. In this case,
The second type of divergence, shown in Figure
10,000 muscle fibers.
highly facilitated conditions, of exciting as many as
large pyramidal cell in the motor cortex capable, under
pathway in its control of skeletal muscles, with a single
through successive orders of neurons in its path. This
This means simply that an input signal spreads
type of divergence is shown in Figure
entirely different purposes.
Two major types of divergence occur and have
gence.
leaving the pool. This phenomenon is called
Neuronal Pools
Divergence of Signals Passing Through
The Nervous System: A. General Principles and Sensory Physiology
580
Unit IX
Often it is important for weak signals entering a neu-
ronal pool to excite far greater numbers of nerve fibers
diver-
An amplifying
46–11A.
to an increasing number of neurons as it passes
type of divergence is characteristic of the corticospinal
46–11B, is divergence into multiple tracts
the signal is transmitted in two directions from the
dorsal columns of the spinal cord takes two courses in
(2) on through the lower regions of the brain to the
into still deeper structures of the thalamus and at the
same time to discrete regions of the cerebral cortex.
Convergence means signals from multiple inputs
A shows
convergence from a single source
minals from a single incoming fiber tract terminate on
neurons are almost never excited by an action poten-
converging on the neuron from multiple terminals
provide enough spatial summation to bring the neuron
Convergence can also result from input signals (exci-
tatory or inhibitory) from multiple sources
B.
spinal cord receive converging signals from (1) periph-
fibers passing from one segment of the cord to
from the interneurons converge on the anterior motor
neurons to control muscle function.
Such convergence allows summation of information
summated effect of all the different types of informa-
Sometimes an incoming signal to a neuronal pool
causes an output excitatory signal going in one direc-
tion and at the same time an inhibitory signal going
tatory signal is transmitted by one set of neurons in the
inhibitory signal is transmitted through a separate set
of neurons to inhibit the muscles on the back of the
leg so that they will not oppose the forward move-
reciprocal inhibition circuit.
mediate inhibitory neuron
a different type of transmitter substance to inhibit the
circuit is also important in preventing overactivity in
many parts of the brain.
Divergence in same tract
A
Divergence in multiple tracts
B
multiple tracts to transmit the signal to separate areas.
“Divergence” in neuronal pathways.
Figure 46–11
A, Divergence within a
pathway to cause “amplification” of the signal. B, Divergence into
Convergence from
single source
A
Convergence from
multiple sources
Source
#3
Source
#2
Source
#1
Source
B
multiple separate sources.
Input fibers from
Multiple input fibers from a single source.
“Convergence” of multiple input fibers onto a single neuron.
Figure 46–12
A,
B,

tions in the circuit. Fatigue beyond a certain critical
it suddenly ceases entirely. The cause of this sudden
beration and then decreases to a critical point, at which
can last for many milliseconds or even minutes. The
lus may last only 1 millisecond or so, and yet the output
from a typical reverberatory circuit. The input stimu-
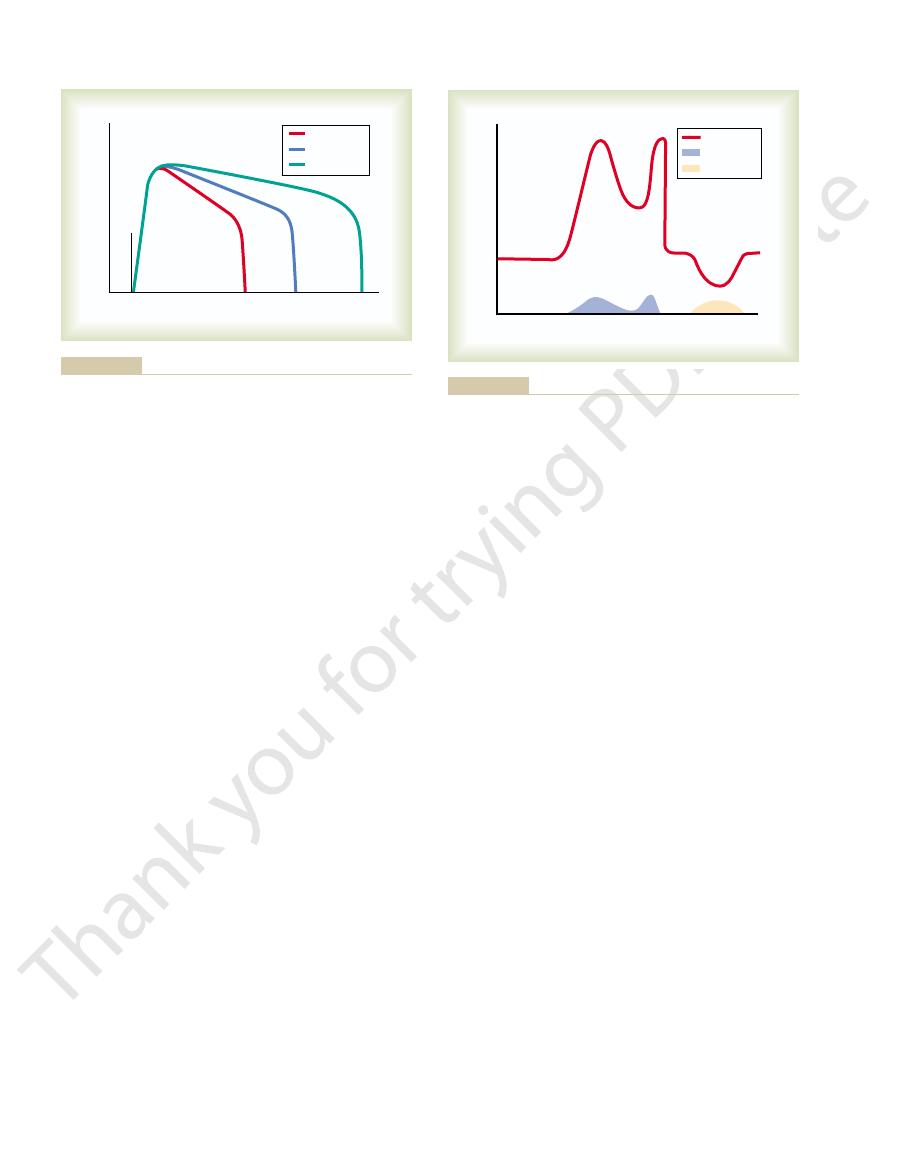
Figure 46–15 shows output signals
Characteristics of Signal Prolongation from a Rever-
weak or strong, depending on how many parallel nerve
a system, the total reverberating signal can be either
cell station, the terminal fibrils spread widely. In such
ways are constituted of many parallel fibers. At each
Figure 46–14
intensity and frequency of reverberation, whereas an
reverberating circuit.A facilitatory signal enhances the
initial discharge and the feedback signal. Figure
feedback circuit, which causes a longer delay between
Figure 46–14
discharging for a protracted time thereafter.
charges, the feedback stimuli could keep the neuron
important one, theoretically, once the neuron dis-
itself. Although this type of circuit probably is not an
case, the output neuron simply sends a collateral nerve
involves only a single neuron. In this
Figure 46–14
are shown in Figure 46–14. The simplest, shown in
for a long time.
once stimulated, the circuit may discharge repetitively
re-excite the input of the same circuit. Consequently,
, or
Reverberatory (Oscillatory) Circuit as a Cause of Signal Prolon-
for many milliseconds.
tic “afterdischarge” mechanism alone, it is possible for
explained in Chapter 45. Thus, as a result of this synap-
mit a continuous train of output impulses, as was
can continue to excite the neuron, causing it to trans-
stances are involved. As long as this potential lasts, it
the neuron and lasts for many milliseconds, especially
neuron, a postsynaptic electrical potential develops in
When excitatory synapses dis-
following.
the incoming signal is over. The most important
, lasting a
afterdischarge
output discharge, called
instances, a signal entering a pool causes a prolonged
relayed through neuronal pools. However, in many
Thus far, we have considered signals that are merely
Prolongation of a Signal by a
Sensory Receptors, Neuronal Circuits for Processing Information
Chapter 46
581
Neuronal Pool—“Afterdischarge”
few milliseconds to as long as many minutes after
mechanisms by which afterdischarge occurs are the
Synaptic Afterdischarge.
charge on the surfaces of dendrites or soma of a
when some of the long-acting synaptic transmitter sub-
a single instantaneous input signal to cause a sustained
signal output (a series of repetitive discharges) lasting
gation.
One of the most important of all circuits in the
entire nervous system is the reverberatory
oscilla-
tory, circuit. Such circuits are caused by positive feed-
back within the neuronal circuit that feeds back to
Several possible varieties of reverberatory circuits
A,
fiber back to its own dendrites or soma to restimulate
B shows a few additional neurons in the
46–14C shows a still more complex system in which
both facilitatory and inhibitory fibers impinge on the
inhibitory signal depresses or stops the reverberation.
D shows that most reverberating path-
fibers are momentarily involved in the reverberation.
beratory Circuit.
figure demonstrates that the intensity of the output
signal usually increases to a high value early in rever-
cessation of reverberation is fatigue of synaptic junc-
level lowers the stimulation of the next neuron in the
circuit below threshold level so that the circuit feed-
back is suddenly broken.
Excitatory synapse
#1
#3
#2
Inhibitory synapse
Input fiber
Excitation
Inhibition
Inhibitory circuit. Neuron 2 is an inhibitory neuron.
Figure 46–13
Input
Output
Input
Output
Input
Input
C
D
B
A
Output
Input
Output
Inhibition
Facilitation
Reverberatory circuits of increasing complexity.
Figure 46–14
rhythmical signals.
dog or the walking movements of any animal, require
throughout life. Other rhythmical signals, such as those
pons. This respiratory rhythmical signal continues
signals—for instance, a rhythmical respiratory signal
berating neuronal pathway.
iris in the eye, and heart rate. That is, the nerve exci-
vascular tone, gut tone, degree of constriction of the
This type of information transmission is used by the
whereas up to this point, the types of information
in signal intensity as well as an increase,
intensity. Note that this carrier wave system allows a
output signal, but they do
information transmission. That is, the excitatory and
Those students who are familiar with radio trans-
greatly increases the output signal, whereas an
of reverberation. Note that an excitatory input signal
a pool of neurons. The pool may be emitting impulses
Figure 46–16 shows a continuous output signal from
increase the output signal, whereas inhibition can
ation is a source of continuous impulses. And excita-
a Means for Transmitting Information.
can decrease the rate of firing to zero.
decreased by inhibitory signals; inhibitory signals often
rons of the spinal cord. The rates at which these cells
of the cerebellum, as well as in most of the interneu-
tinually. This occurs especially in many of the neurons
potential rises above a certain threshold level. The
Neurons, like other excitable tissues, discharge
ity.
beratory signals.
two mechanisms can cause this effect: (1) continuous
ously, even without excitatory input signals. At least
Neuronal Circuits
Continuous Signal Output from Some
in Figure 46–18).
brain that inhibit or facilitate the circuit. Almost these
The duration of the total signal before cessation can
The Nervous System: A. General Principles and Sensory Physiology
582
Unit IX
also be controlled by signals from other parts of the
exact patterns of output signals are recorded from the
motor nerves exciting a muscle involved in a flexor
reflex after pain stimulation of the foot (as shown later
Some neuronal circuits emit output signals continu-
intrinsic neuronal discharge and (2) continuous rever-
Continuous Discharge Caused by Intrinsic Neuronal Excitabil-
repetitively if their level of excitatory membrane
membrane potentials of many neurons even normally
are high enough to cause them to emit impulses con-
emit impulses can be increased by excitatory signals or
Continuous Signals Emitted from Reverberating Circuits as
A reverberating
circuit that does not fatigue enough to stop reverber-
tory impulses entering the reverberating pool can
decrease or even extinguish the signal.
because of intrinsic neuronal excitability or as a result
inhibitory input signal greatly decreases the output.
mitters will recognize this to be a carrier wave type of
inhibitory control signals are not the cause of the
control its changing level of
decrease
transmission we have discussed have been mainly pos-
itive information rather than negative information.
autonomic nervous system to control such functions as
tatory signal to each of these can be either increased
or decreased by accessory input signals into the rever-
Rhythmical Signal Output
Many neuronal circuits emit rhythmical output
originates in the respiratory centers of the medulla and
that cause scratching movements by the hind leg of a
input stimuli into the respective circuits to initiate the
All or almost all rhythmical signals that have been
studied experimentally have been found to result from
reverberating circuits or a succession of sequential
reverberating circuits that feed excitatory or inhibitory
signals in a circular pathway from one neuronal pool
to the next.
Output pulse rate
Time
Inhibited
Input stimulus
Facilitated
Normal
after a single input stimulus, showing the effects of facilitation and
Typical pattern of the output signal from a reverberatory circuit
Figure 46–15
inhibition.
Impulses per second
Time
Output
Excitation
Inhibition
of excitatory or inhibitory input signals.
intrinsically discharging neurons. This figure also shows the effect
Continuous output from either a reverberating circuit or a pool of
Figure 46–16
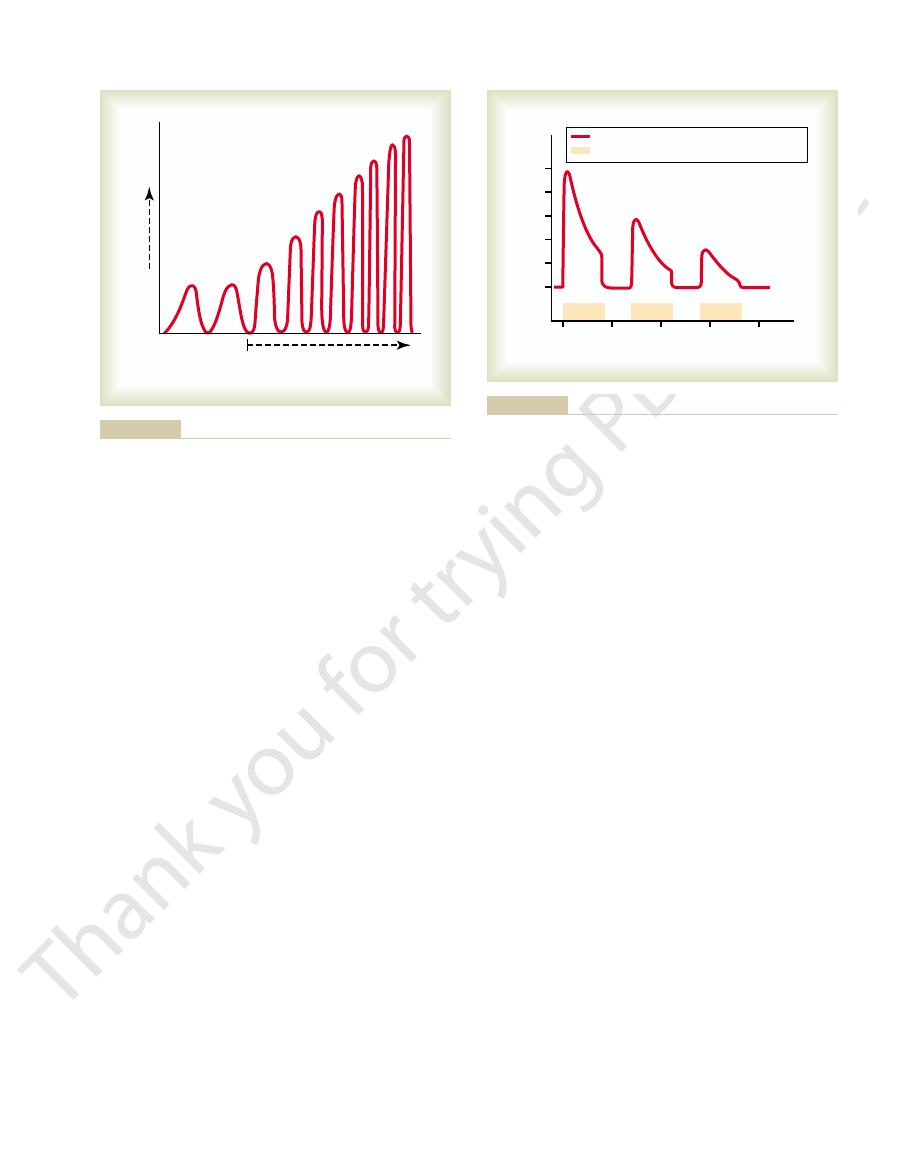
of fatigue to other pathways in the brain. Those that
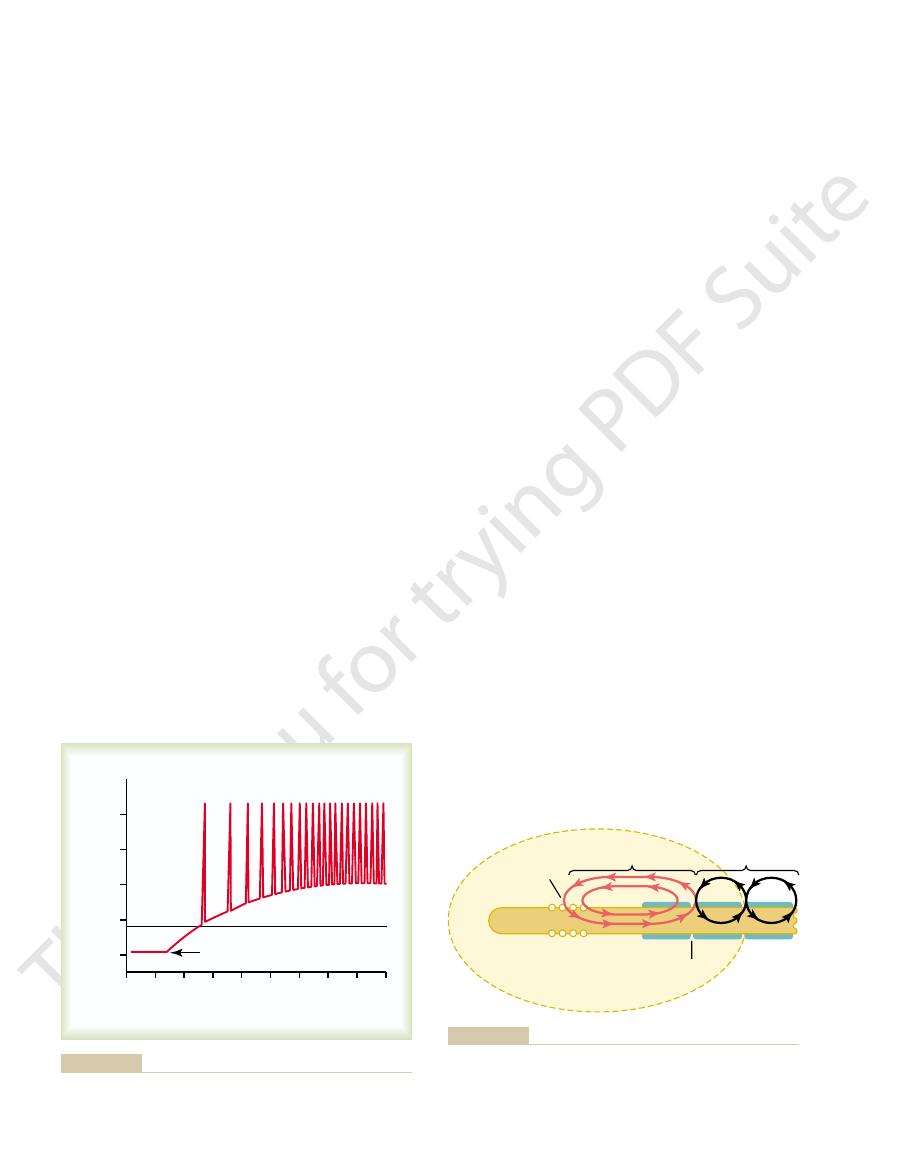
Automatic Short-Term Adjustment of Pathway Sensitivity by the
subsequent reflex response.
successive flexor reflexes, the less the intensity of the
circuit. Furthermore, the shorter the interval between
that is, its strength diminishes; much of this effect is
strength of contraction progressively “decrements”—
the footpad of the paw. Note in each record that the
Figure 46–18 shows three successive records of a flexor
Synaptic Fatigue as a Means for
brain—for instance, many of the basal ganglia exert
overly excited; and (2) some neuronal pools that exert
Two types of inhibitory circuits in widespread areas
Stabilizing Nervous System Function
Inhibitory Circuits as a Mechanism for
fatigue of synapses.
central nervous system: (1) inhibitory circuits and (2)
from happening all the time? The answer lies mainly
epileptic seizures.
signals could be transmitted. Such an effect occurs in
mation but, nevertheless, would be consuming the cir-
parts. If this should occur, the brain would be inun-
would set off a continuous cycle of re-excitation of all
finally the signal re-excites the first part, it is clear that
second the third, the third the fourth, and so on until
serious problem. If the first part excites the second, the
or indirectly with every other part, and this creates a
of Neuronal Circuits
progressively.
deficiency, both the frequency and the amplitude of
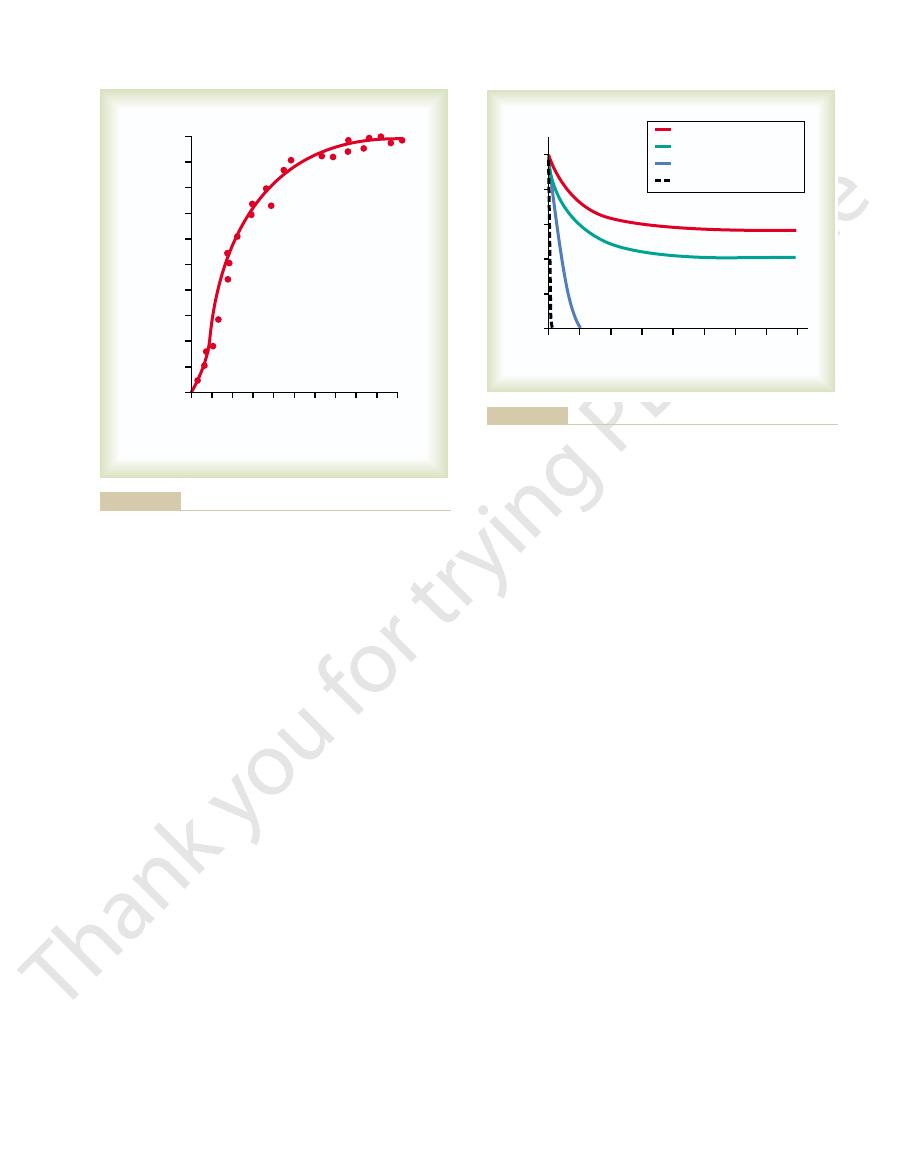
When the carotid body is stimulated by arterial oxygen
the respiratory signal output in the phrenic nerve.
output. Figure 46–17, for instance, shows changes in
Sensory Receptors, Neuronal Circuits for Processing Information
Chapter 46
583
Excitatory or inhibitory signals can also increase
or decrease the amplitude of the rhythmical signal
the respiratory rhythmical output signal increase
Instability and Stability
Almost every part of the brain connects either directly
an excitatory signal entering any part of the brain
dated by a mass of uncontrolled reverberating
signals—signals that would be transmitting no infor-
cuits of the brain so that none of the informational
widespread areas of the brain during
How does the central nervous system prevent this
in two basic mechanisms that function throughout the
of the brain help prevent excessive spread of signals:
(1) inhibitory feedback circuits that return from the
termini of pathways back to the initial excitatory
neurons of the same pathways—these circuits occur in
virtually all sensory nervous pathways and inhibit
either the input neurons or the intermediate neurons
in the sensory pathway when the termini become
gross inhibitory control over widespread areas of the
inhibitory influences throughout the muscle control
system.
Stabilizing the Nervous System
Synaptic fatigue means simply that synaptic transmis-
sion becomes progressively weaker the more pro-
longed and more intense the period of excitation.
reflex elicited in an animal caused by inflicting pain in
caused by fatigue of synapses in the flexor reflex
Fatigue Mechanism.
Now let us apply this phenomenon
Phrenic nerve output
Increasing carotid
body stimulation
respiration.
quency of the phrenic nerve signal to the diaphragm to increase
tion of the carotid body increases both the intensity and the fre-
respiratory center, showing that progressively increasing stimula-
The rhythmical output of summated nerve impulses from the
Figure 46–17
0
15
30
45
60
Flexor muscle contraction force (g)
Seconds
0
10
20
30
40
50
Flexor reflexes–decremental responses
Stimulus
the reflex pathway.
Successive flexor reflexes showing fatigue of conduction through
Figure 46–18

81:299, 2001.
adaptations mediating opioid dependence. Physiol Rev
Williams JT, Christie MJ, Manzoni O: Cellular and synaptic
synapse formation. Curr Opin Neurobiol 14:83, 2004.
Shen K: Molecular mechanisms of target specificity during
Physiol Rev 82:875, 2002.
Schwartz EA: Transport-mediated synapses in the retina.
Neurophysiol 90:1363, 2003.
mitter transporters: not just for reuptake anymore. J
Richerson GB, Wu Y: Dynamic equilibrium of neurotrans-
glycine receptors and taurine. Neuron 41:839, 2004.
Renteria RC, Johnson J, Copenhagen DR: Need rods? Get
mic behavior. Annu Rev Physiol 62:723, 2000.
Pearson KG: Neural adaptation in the generation of rhyth-
tonic neurons. Neuron 35:600, 2002.
Palmer MJ, von Gersdorff H: Phasic transmitter release from
and taste receptors. Nat Rev Neurosci 5:263, 2004.
Mombaerts P: Genes and ligands for odorant, vomeronasal
bution? Acta Physiol Scand 177:227, 2003.
thetic ganglia—modulation, integration or simply distri-
McLachlan EM: Transmission of signals through sympa-
tic plasticity? Trends Neurosci 27:62, 2004.
Krupa B, Liu G: Does the fusion pore contribute to synap-
Science, 4th ed. New York: McGraw-Hill, 2000.
Kandel ER, Schwartz JH, Jessell TM: Principles of Neural
bridge, MA: MIT Press, 1998.
Ivry RB, Robertson LC: The Two Sides of Perception. Cam-
duction in living cells. Physiol Rev 81:685, 2001.
Hamill OP, Martinac B: Molecular basis of mechanotrans-
Biobehav Rev 27:729, 2004.
Gebhart GF: Descending modulation of pain. Neurosci
muscle fatigue. Physiol Rev 81:1725, 2001.
Gandevia SC: Spinal and supraspinal factors in human
tion in vertebrate photoreceptors. Physiol Rev 81:117,
Fain GL, Matthews HR, Cornwall MC, Koutalos Y: Adapta-
Basar E: Brain Function and Oscillations. Berlin: Springer,
Baev KV: Biological Neural Networks. Boston: Birkhauser,
Nat Neurosci 7:446, 2004.
Buzsaki G: Large-scale recording of neuronal ensembles.
or other nervous disorders. But fortunately, the auto-
chotic disturbances, hallucinations, mental tension,
expect almost continual muscle cramps, seizures, psy-
of these circuits were abnormally high; one might then
level required for proper function. Think for a moment
anisms for adjusting synaptic sensitivity, continually
regulation of receptors, as well as other control mech-
tic membrane.
combine with the receptor proteins, many of these
synaptic membrane. However, when the synapses are
overactivity. The mechanism for this is the following:
tivity, and downregulating the receptors when there is
The
Long-Term Changes in Synaptic Sensitivity Caused by Automatic
cuits. These help to keep the circuits operating in a
increase. Thus, fatigue and recovery from fatigue con-
underused become rested, and their sensitivities
sensitivities decrease. Conversely, those that are
are overused usually become fatigued, so that their
The Nervous System: A. General Principles and Sensory Physiology
584
Unit IX
stitute an important short-term means of moderating
the sensitivities of the different nervous system cir-
range of sensitivity that allows effective function.
Downregulation or Upregulation of Synaptic Receptors.
long-term sensitivities of synapses can be changed
tremendously by upregulating the number of receptor
proteins at the synaptic sites when there is underac-
Receptor proteins are being formed constantly by the
endoplasmic reticular–Golgi apparatus system and are
constantly being inserted into the receptor neuron
overused so that excesses of transmitter substance
receptors are inactivated and removed from the synap-
It is indeed fortunate that upregulation and down-
adjust the sensitivity in each circuit to almost the exact
how serious it would be if the sensitivities of only a few
matic controls normally readjust the sensitivities of the
circuits back to controllable ranges of reactivity any
time the circuits begin to be too active or too
depressed.
References
1998.
1998.
2001.