The cerebellum also aids the cerebral cortex in planning the next sequential
activation of specific muscles.
not compare favorably, then instantaneous subconscious corrective signals are
information with the movements intended by the motor system. If the two do
rate of movement, forces acting on it, and so forth. The cerebellum then
receives continuous sensory information from the peripheral parts of the body,
sequence of muscle contractions from the brain motor control areas; it also
The cerebellum receives continuously updated information about the desired
the motor signals directed by the cerebral motor cortex and other parts of the
s motor activities while they are being executed so that they will conform to
ability to cause muscle contraction? The answer is that it helps to
ties even though its loss causes paralysis of no muscles.
cular activities such as running, typing, playing the piano, and even talking. Loss
become highly abnormal. The cerebellum is especially vital during rapid mus-
ment. Removal of the cerebellum, however, does cause body movements to
of the brain, principally because electrical excitation of the cerebel-
The cerebellum, illustrated in Figures 56–1 and 56–2, has long been called a
Cerebellum and Its Motor Functions
activity.
achieving specific complicated motor goals. This chapter explains the basic
movements, and sequencing of multiple successive and parallel movements for
, controlling relative intensities of the separate movements, directions of
basal ganglia help to plan and control complex patterns of muscle move-
The
interplay between agonist and antagonist muscle groups.
when the muscle load changes, as well as controlling necessary instantaneous
movement to the next. It also helps to control intensity of muscle contraction
timing of motor activities and in rapid, smooth progression from one muscle
Basically, the
they always function in associa-
themselves. Instead,
. Yet
They are the
stimulate muscle contraction, two other brain struc-
C
H
A
P
T
E
R
5
6
698
Contributions of the Cerebellum
and Basal Ganglia to Overall
Motor Control
Aside from the areas in the cerebral cortex that
tures are also essential for normal motor function.
cerebellum and the basal ganglia
neither of these two can control muscle function by
tion with other systems of motor control.
cerebellum plays major roles in the
ment
mechanisms of function of the cerebellum and basal ganglia and discusses the
overall brain mechanisms for achieving intricate coordination of total motor
silent area
lum does not cause any conscious sensation and rarely causes any motor move-
of this area of the brain can cause almost total incoordination of these activi-
But how is it that the cerebellum can be so important when it has no direct
sequence
the motor activities and also monitors and makes corrective adjustments in the
body’
brain.
giving sequential changes in the status of each part of the body—its position,
com-
pares the actual movements as depicted by the peripheral sensory feedback
transmitted back into the motor system to increase or decrease the levels of
movement a fraction of a second in advance while the current movement is still
bral cortex and brain stem. In turn, they send motor
facial regions lie in the intermediate zones. These topo-
vermis part of the cerebellum, whereas the limbs and
tions. Note that the axial portions of the body lie in the
cerebellum. Figure 56–3 shows two such representa-
sentations of the different parts of the body, so also is
sensory cortex, motor cortex, basal ganglia, red nuclei,
Topographical Representation of the Body in the Vermis and Inter-
nate, as we discuss more fully later.
motor movements. Without this lateral zone, most dis-
The lateral zone of the hemisphere operates at a
hands and fingers and feet and toes.
portions of the upper and lower limbs, especially the
The intermediate zone of the hemisphere is con-
zone.
intermediate zone
, and each of these hemi-
To each side of the vermis is a large, laterally pro-
neck
cerebellum by shallow grooves. In this area, most cere-
, separated from the remainder of the
longitudinal axis, as demonstrated in Figure 56–2, which
From a functional point of view, the anterior and
controlling body equilibrium, as discussed in Chapter
oldest of all portions of the cerebellum; it developed
The flocculonodular lobe is the
flocculonodular lobe.
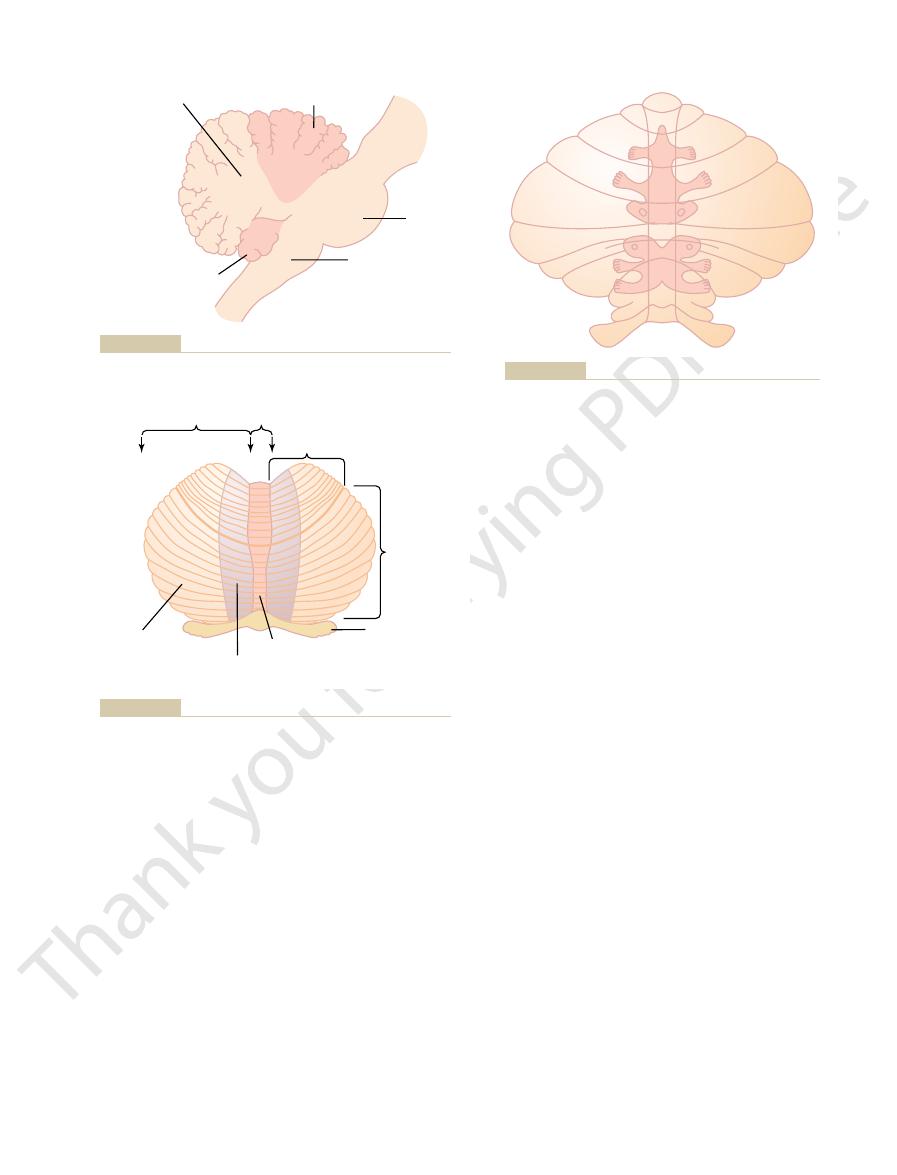
, and (3) the
, (2) the
by two deep fissures, as shown in Figures 56–1 and 56–2:
Anatomically, the cerebellum is divided into three lobes
of the Cerebellum
Anatomical Functional Areas
intended movements
priate cerebellar neurons, thus bringing subsequent
changes occur in the excitability of appro-
To do this,
to make a stronger or weaker movement the next time.
occur exactly as intended, the cerebellar circuit learns
learns by its mistakes—that is, if a movement does not
smoothly from one movement to the next. Also, it
being executed, thus helping the person to progress
Chapter 56
Contributions of the Cerebellum and Basal Ganglia to Overall Motor Control
699
muscle contractions into better correspondence with the
.
(1) the anterior lobe
posterior lobe
along with (and functions with) the vestibular system in
55.
Longitudinal Functional Divisions of the Anterior and Posterior
Lobes.
posterior lobes are organized not by lobes but along the
shows a posterior view of the human cerebellum after
the lower end of the posterior cerebellum has been
rolled downward from its normally hidden position.
Note down the center of the cerebellum a narrow band
called the vermis
bellar control functions for muscle movements of the
axial body,
, shoulders, and hips are located.
truding cerebellar hemisphere
spheres is divided into an
and a lateral
cerned with controlling muscle contractions in the distal
much more remote level because this area joins with the
cerebral cortex in the overall planning of sequential
crete motor activities of the body lose their appropriate
timing and sequencing and therefore become incoordi-
mediate Zones.
In the same manner that the cerebral
and reticular formation all have topographical repre-
this true for the vermis and intermediate zones of the
graphical representations receive afferent nerve signals
from all the respective parts of the body as well as from
corresponding topographical motor areas in the cere-
Pons
Anterior lobe
Posterior lobe
Medulla
Flocculonodular
lobe
Anatomical lobes of the cerebellum as seen from the lateral side.
Figure 56–1
Hemisphere
Vermis
Vermis
Intermediate zone
of hemisphere
Anterior
lobe
Posterior
lobe
Lateral zone
of hemisphere
Flocculonodular
lobe
outward to flatten the surface.
rior view, with the inferiormost portion of the cerebellum rolled
Functional parts of the cerebellum as seen from the posteroinfe-
Figure 56–2
Somatosensory projection areas in the cerebellar cortex.
Figure 56–3
Thus, this ventral fiber pathway tells the cerebellum
the internal motor pattern generators in the cord itself.
less information from the peripheral receptors. Instead,
Conversely, the ventral spinocerebellar tracts receive
surfaces of the body.
of the parts of the body, and (4) forces acting on the
muscle tendons, (3) positions and rates of movement
of (1) muscle contraction, (2) degree of tension on the
receptors of the skin, and joint receptors. All these
the body, such as Golgi tendon organs, large tactile
The signals transmitted in the dorsal spinocerebellar
through the superior cerebellar peduncle, but it termi-
as its origin. The ventral tract enters the cerebellum
The
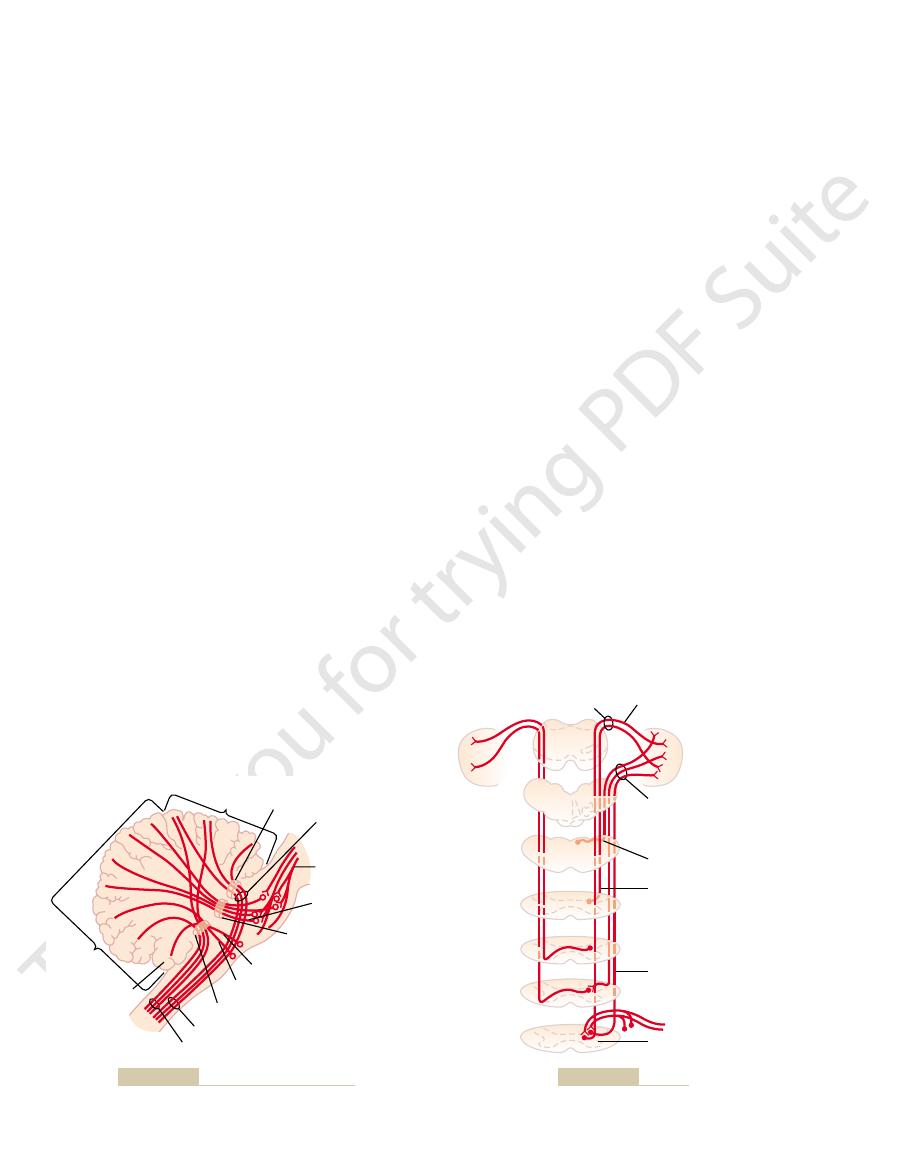
tracts are shown in Figure 56–5: the
cord and two ventrally. The two most important of these
on each side, two of which are located dorsally in the
The cerebellum also
Afferent Pathways from the Periphery.
, which originate in different portions of the brain
of the cerebellum; and (3)
, some of which
; (2)
spinal cord
, and
, widespread areas of the
, which passes from the
each side of the brain stem; they include (1) an exten-
In addition, important afferent tracts originate in
the opposite side of the brain from the cerebral areas.
, which originates in
corticopontocerebellar pathway
56–4. An extensive and important afferent pathway is
input pathways to the cerebellum are shown in Figure
The basic
56–2 and 56–3. Each fold is called a folium. Lying deep
long, with the folds lying crosswise, as shown in Figures
sheet, about 17 centimeters wide by 120 centimeters
The human cerebellar cortex is actually a large folded
Neuronal Circuit of the Cerebellum
important roles in planning and coordinating the body’s
association areas of the parietal cortex. It is believed
cortex, especially from the premotor areas of the frontal
of the body. These areas of the cerebellum receive their
of the cerebral motor cortex, as well as to topographi-
The Nervous System: C. Motor and Integrative Neurophysiology
700
Unit XI
signals back to the same respective topographical areas
cal areas of the red nucleus and reticular formation in
the brain stem.
Note that the large lateral portions of the cerebellar
hemispheres do not have topographical representations
input signals almost exclusively from the cerebral
cortex and from the somatosensory and other sensory
that this connectivity with the cerebral cortex allows the
lateral portions of the cerebellar hemispheres to play
rapid sequential muscular activities that occur one after
another within fractions of a second.
beneath the folded mass of cerebellar cortex are deep
cerebellar nuclei.
Input Pathways to the Cerebellum
Afferent Pathways from Other Parts of the Brain.
the
the cerebral motor and premotor cortices and also in the
cerebral somatosensory cortex. It passes by way of the
pontile nuclei and pontocerebellar tracts mainly to
the lateral divisions of the cerebellar hemispheres on
sive olivocerebellar tract
inferior
olive to all parts of the cerebellum and is excited in the
olive by fibers from the cerebral motor cortex, basal
ganglia
reticular formation
vestibulocerebellar fibers
originate in the vestibular apparatus itself and others
from the brain stem vestibular nuclei—almost all of
these terminate in the flocculonodular lobe and fastigial
nucleus
reticulocerebellar
fibers
stem reticular formation and terminate in the midline
cerebellar areas (mainly in the vermis).
receives important sensory signals directly from the
peripheral parts of the body mainly through four tracts
dorsal spinocere-
bellar tract and the ventral spinocerebellar tract.
dorsal tract enters the cerebellum through the inferior
cerebellar peduncle and terminates in the vermis and
intermediate zones of the cerebellum on the same side
nates in both sides of the cerebellum.
tracts come mainly from the muscle spindles and to a
lesser extent from other somatic receptors throughout
signals apprise the cerebellum of the momentary status
they are excited mainly by motor signals arriving in the
anterior horns of the spinal cord from (1) the brain
through the corticospinal and rubrospinal tracts and (2)
Superior cerebellar
peduncle
Ventral
spinocerebellar
tract
Cerebropontile
tract
Pontocerebellar
tract
Middle cerebellar
peduncle
Vestibulocerebellar tract
Olivocerebellar and
reticulocerebellar tract
Inferior cerebellar peduncle
Ventral spinocerebellar tract
Dorsal spinocerebellar tract
Flocculonodular
lobe
Anterior
lobe
Posterior
lobe
tracts to the cerebellum.
afferent
Figure 56–4
Principal
Superior cerebellar peduncle
Dorsal spinocerebellar tract
Clark’s cells
Spinal cord
Ventral spinocerebellar tract
Dorsal external arcuate fiber
Medulla oblongata
Inferior cerebellar peduncle
Ventral spinocerebellar tract
Cerebellum
Spinocerebellar tracts.
Figure 56–5

. There is one climbing fiber for
The climbing fibers
two types, one called the
The afferent inputs to the cerebellum are mainly of
bellum from the brain or the periphery. The inhibitory
influences. The excitatory influences arise from direct
. This cell
30 million times in the cerebellum. The output from
functional unit, which is repeated with little variation
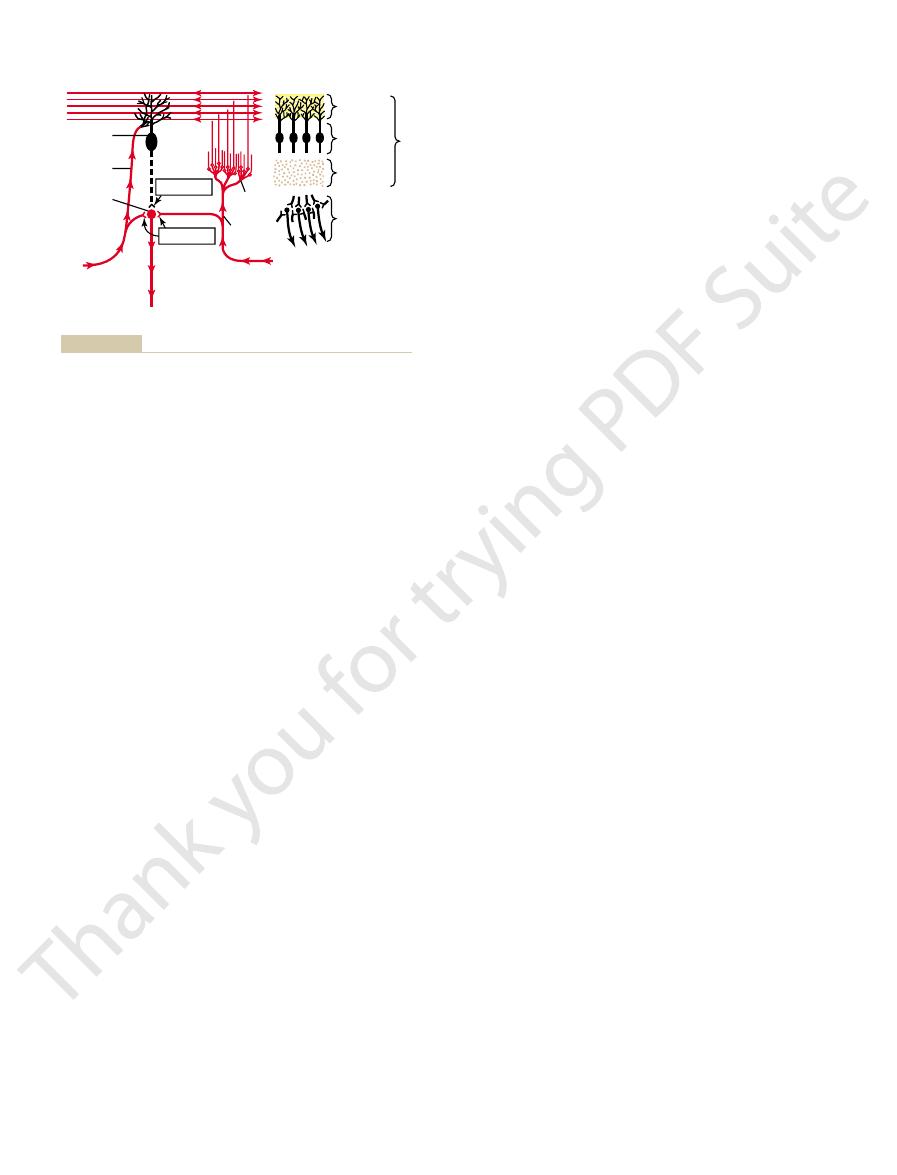
left half of Figure 56–7 is the neuronal circuit of the
bellar mass, are the deep cerebellar nuclei that send
Beneath these cortical layers, in the center of the cere-
granule cell layer
, and
Purkinje cell layer
lar layer
layers of the cerebellar cortex are shown: the
To the top and right in Figure 56–7, the three major
Figure 56–7. This functional unit centers on a single,
functional units, one of which is shown to the left in
The cerebellum has about 30 million nearly identical
Functional Unit of the Cerebellar Cortex—
thalamus, and, finally, to the cerebral cortex. This
then passes to the dentate nucleus, next to the
3. A pathway that begins in the cerebellar cortex of
limbs, especially in the hands, fingers, and thumbs.
brain stem. This complex circuit helps to coordinate
thalamus and then to (4) the cerebral cortex, to (5)
2. A pathway that originates in (1) the intermediate
postural attitudes of the body. It was discussed in
control equilibrium, and also in association with the
This circuit
1. A pathway that originates in the
leading out of the cerebellum is shown in Figure 56–6
The general plan of the major efferent pathways
inhibitory signals. From the deep nuclei, output signals
Thus, all input signals that enter the cerebellum even-
output signal to the deep nucleus.
Then, a fraction of a second later, the cerebellar cortex
area of the cerebellar cortex overlying the deep nucleus.
it divides and goes in two directions: (1) directly to one
receive signals from two sources: (1) the cerebellar
the flocculonodular lobe.) All the deep cerebellar nuclei
(The
, and
these areas to the cerebellum. Thus, the cerebellum con-
olivary nucleus. Then signals are relayed from both of
spino-olivary pathway
spinoreticular pathway
to the cerebellum. Likewise, signals are transmitted up
In addition to signals from the spinocerebellar tracts,
in peripheral muscle actions.
system. This extremely rapid conduction is important
at velocities up to 120 m/sec, which is the most rapid
The spinocerebellar pathways can transmit impulses
horn motor drive.
efference copy
Chapter 56
Contributions of the Cerebellum and Basal Ganglia to Overall Motor Control
701
which motor signals have arrived at the anterior horns;
this feedback is called the
of the anterior
conduction in any pathway in the central nervous
for instantaneous apprisal of the cerebellum of changes
signals are transmitted into the cerebellum from the
body periphery through the spinal dorsal columns to the
dorsal column nuclei of the medulla and then relayed
the spinal cord through the
to
the reticular formation of the brain stem and also
through the
to the inferior
tinually collects information about the movements and
positions of all parts of the body even though it is oper-
ating at a subconscious level.
Output Signals from the Cerebellum
Deep Cerebellar Nuclei and the Efferent Pathways.
Located
deep in the cerebellar mass on each side are three deep
cerebellar nuclei—the dentate, interposed
fastigial.
vestibular nuclei in the medulla also function in
some respects as if they were deep cerebellar nuclei
because of their direct connections with the cortex of
cortex and (2) the deep sensory afferent tracts to the
cerebellum.
Each time an input signal arrives in the cerebellum,
of the cerebellar deep nuclei and (2) to a corresponding
relays an inhibitory
tually end in the deep nuclei in the form of initial exci-
tatory signals followed a fraction of a second later by
leave the cerebellum and are distributed to other parts
of the brain.
and consists of the following:
midline structures
of the cerebellum (the vermis) and then passes
through the fastigial nuclei into the medullary and
pontile regions of the brain stem.
functions in close association with the equilibrium
apparatus and brain stem vestibular nuclei to
reticular formation of the brain stem to control the
detail in Chapter 55 in relation to equilibrium.
zone of the cerebellar hemisphere and then passes
through (2) the interposed nucleus to (3) the
ventrolateral and ventroanterior nuclei of the
several midline structures of the thalamus and then
to (6) the basal ganglia and (7) the red nucleus and
reticular formation of the upper portion of the
mainly the reciprocal contractions of agonist and
antagonist muscles in the peripheral portions of the
the lateral zone of the cerebellar hemisphere and
ventrolateral and ventroanterior nuclei of the
pathway plays an important role in helping
coordinate sequential motor activities initiated by
the cerebral cortex.
The Purkinje Cell and the Deep Nuclear Cell
very large Purkinje cell (30 million of which are in
the cerebellar cortex) and on a corresponding deep
nuclear cell.
molecu-
,
.
output signals to other parts of the nervous system.
Neuronal Circuit of the Functional Unit.
Also shown in the
the functional unit is from a deep nuclear cell
is continually under both excitatory and inhibitory
connections with afferent fibers that enter the cere-
influence arises entirely from the Purkinje cell in the
cortex of the cerebellum.
climbing fiber type and the
other called the mossy fiber type.
all originate from the inferior
olives of the medulla
Fastigioreticular tract
Cerebellothalamocortical
tract
Dentate
Superior cerebellar
peduncle
Reticulum of
mesencephalon
Red nucleus
To thalamus
Fastigioreticular tract
Fastigial nucleus
Paleocerebellum
tracts from the cerebellum.
efferent
Figure 56–6
Principal
are not fully known, one can speculate from the basic
signals to the antagonists. Although the exact details
on approaching termination of the movement, the
antagonist muscles at the onset of a movement. Then
The typical function of the cerebellum is to help
Signals from the Cerebellum
Turn-On/Turn-Off and Turn-Off/Turn-On Output
of adjacent Purkinje cells, thus sharpening
right angles across the parallel fibers and cause
parallel fibers. These cells in turn send their axons at
lar cortex, lying among and stimulated by the small
with short axons. Both the basket cells and the stellate
. These are inhibitory cells
deep nuclear cells, granule cells, and Purkinje cells, two
Other Inhibitory Cells in the Cerebellum.
occur.
mark. Otherwise, oscillation of the movement would
. That is, when the motor system is excited,
signal resembles a “delay-line” negative feedback
of a second by an inhibitory signal. This inhibitory
ment, but this is followed within another small fraction
circuit arrive. In this way, there is first a rapid excita-
cell excitation. Then, another few milliseconds later,
In execution of a rapid motor movement, the
tation, so that, under quiet conditions, output from the
the Purkinje cells inhibit them. Normally, the balance
fibers excites them. By contrast, signals arriving from
56–7, one should note that direct stimulation of the
Referring again to the circuit of Figure
more, the output activity of both these cells can be
the deep nuclear cells at much higher rates. Further-
at about 50 to 100 action potentials per second, and
both of them fire continuously; the Purkinje cell fires
, rather than the prolonged complex action
excite the Purkinje cell. Furthermore, activation
synaptic connections are weak, so that large numbers
The mossy fiber input to the Purkinje cell is quite
project and 80,000 to 200,000 of the parallel fibers
granule cells for every 1 Purkinje cell. It is into this
because there are some 500 to 1000
lel to the folia. There are many millions of these
bellar cortex. Here the axons divide into two branches
small axons, less than 1 micrometer in diameter, up to
. In turn, the granule cells send very, very
send collaterals to excite the deep nuclear cells. Then
brain, brain stem, and spinal cord. These fibers also
the cerebellum from multiple sources: from the higher
The mossy fibers are all the other fibers that enter
secondary spikes. This action potential is called the
Purkinje cell with which it connects, beginning with
second), peculiar type of action potential in each
in it will always cause a single, prolonged (up to 1
and dendrites of each Purkinje cell. This climbing
several deep nuclear cells, the climbing fiber continues
about 5 to 10 Purkinje cells. After sending branches to
The Nervous System: C. Motor and Integrative Neurophysiology
702
Unit XI
all the way to the outer layers of the cerebellar cortex,
where it makes about 300 synapses with the soma
fiber is distinguished by the fact that a single impulse
a strong spike and followed by a trail of weakening
complex spike.
they proceed to the granule cell layer of the cortex,
where they too synapse with hundreds to thousands of
granule cells
the molecular layer on the outer surface of the cere-
that extend 1 to 2 millimeters in each direction paral-
par-
allel nerve fibers
molecular layer that the dendrites of the Purkinje cells
synapse with each Purkinje cell.
different from the climbing fiber input because their
of mossy fibers must be stimulated simultaneously to
usually takes the form of a much weaker short-
duration Purkinje cell action potential called a simple
spike
potential caused by climbing fiber input.
Purkinje Cells and Deep Nuclear Cells Fire Continuously Under
Normal Resting Conditions.
One characteristic of both
Purkinje cells and deep nuclear cells is that normally
modulated upward or downward.
Balance Between Excitation and Inhibition at the Deep Cere-
bellar Nuclei.
deep nuclear cells by both the climbing and the mossy
between these two effects is slightly in favor of exci-
deep nuclear cell remains relatively constant at a mod-
erate level of continuous stimulation.
initiating signal from the cerebral motor cortex or
brain stem at first greatly increases deep nuclear
feedback inhibitory signals from the Purkinje cell
tory signal sent by the deep nuclear cells into the
motor output pathway to enhance the motor move-
signal of the type that is effective in providing
damping
a negative feedback signal occurs after a short delay
to stop the muscle movement from overshooting its
In addition to the
other types of neurons are located in the cerebellum:
basket cells and stellate cells
cells are located in the molecular layer of the cerebel-
lateral
inhibition
the signal in the same manner that lateral inhibition
sharpens contrast of signals in many other neuronal
circuits of the nervous system.
provide rapid turn-on signals for the agonist muscles
and simultaneous reciprocal turn-off signals for the
cerebellum is mainly responsible for timing and exe-
cuting the turn-off signals to the agonists and turn-on
Purkinje
cell
Climbing
fiber
Deep
nuclear
cell
Input
(inferior olive)
Output
Excitation
Inhibition
Granule
cells
Cortex
Input
(all other afferents)
Mossy
fiber
Molecular
layer
Purkinje
cell layer
Granule
cell layer
Deep
nuclei
bellar cortex with its three layers.
the physical relationship of the deep cerebellar nuclei to the cere-
cell (an inhibitory neuron) shown in black. To the right is shown
cerebellum, with excitatory neurons shown in red and the Purkinje
The left side of this figure shows the basic neuronal circuit of the
Figure 56–7

. This consists principally
1. The
motor control functions at three levels, as follows:
The nervous system uses the cerebellum to coordinate
Function of the Cerebellum in Overall
cerebellum to cause further change.
fibers no longer need to send “error” signals to the
perfection. When this has been achieved, the climbing
cerebellum, is believed to make the timing and other
along with other possible “learning” functions of the
cells. Over a period of time, this change in sensitivity,
the intended movement. And the climbing fiber signals
for the first time, feedback signals from the muscle and
ing spikes. When a person performs a new movement
During this time, the Purkinje cell fires with one initial
tree of the Purkinje cell, lasting for up to a second.
about once per second. But each time they do fire, they
Under resting conditions, the climbing fibers fire
granule cell excitation becomes altered. Furthermore,
sively adapt during the training process. Especially,
answer is not known, although it is known that sensi-
How do these adjustments come about? The exact
other times requiring hundreds of movements.
ments before the desired result is achieved, but at
more precise, sometimes requiring only a few move-
times, the individual events become progressively
movement. But after the act has been performed many
the end of contraction, and the timing of these are
at the onset of contraction, the degree or inhibition at
ically, when a person first performs a new motor act,
contractions, must be learned by the cerebellum. Typ-
and offset of muscle contractions, as well as timing of
The degree to which the cerebellum supports onset
Errors—Role of the Climbing Fibers
The Purkinje Cells “Learn” to Correct Motor
antagonist muscles, and controlling the timing as well.
on and turn-off signals, controlling the agonist and
They are presented here especially to illustrate ways
are still to be determined; they, too, could play roles in
besides Purkinje cells. The functions of some of these
muscles. But we must remember, too, that the cere-
ment, mirroring whatever occurs in the agonist
the cord can initiate. Therefore, these circuits are part
nist muscles. Most important, remember that through-
Thus, one can see how the complete cerebellar
time.
fore, this helps
that had originally turned on the movement. There-
But once the Purkinje cell is excited, it in turn sends a
have diameters of only a fraction of a millimeter. Also,
fibers of the cerebellar cortical molecular layer, which
nerve fibers in the nervous system: that is, the parallel
through some of the smallest, slowest-conducting
the deep nuclear cells. This pathway passes
fibers, to the Purkinje cells. The Purkinje cells in turn
cerebellar cortex and eventually, by way of “parallel”
Now, what causes the turn-off signal for the agonist
missing. This cerebellar support makes the turn-on
cerebellum, the secondary extra supportive signal is
when the cerebellum is intact, but in the absence of the
and the cerebellar signals. This is the normal effect
sequence, the turn-on signal, after a few milliseconds,
already been begun by the cerebral cortex. As a con-
stem, to support the muscle contraction signal that had
into the cerebral corticospinal motor system, either by
nuclei; this instantly sends an excitatory signal back
the pontile mossy fibers into the cerebellum. One
At the same time, parallel signals are sent by way of
These signals pass through noncerebellar brain stem
as follows.
cerebellar circuit of Figure 56–7 how this might work,
Chapter 56
Contributions of the Cerebellum and Basal Ganglia to Overall Motor Control
703
Let us suppose that the turn-on/turn-off pattern of
agonist/antagonist contraction at the onset of move-
ment begins with signals from the cerebral cortex.
and cord pathways directly to the agonist muscle to
begin the initial contraction.
branch of each mossy fiber goes directly to deep
nuclear cells in the dentate or other deep cerebellar
way of return signals through the thalamus to the cere-
bral cortex or by way of neuronal circuitry in the brain
becomes even more powerful than it was at the start
because it becomes the sum of both the cortical
muscle contraction much stronger than it would be if
the cerebellum did not exist.
muscles at the termination of the movement? Remem-
ber that all mossy fibers have a second branch that
transmits signals by way of the granule cells to the
inhibit
the signals from these fibers are weak so that they
require a finite period of time to build up enough exci-
tation in the dendrites of the Purkinje cell to excite it.
strong inhibitory signal to the same deep nuclear cell
to turn off the movement after a short
circuit could cause a rapid turn-on agonist muscle con-
traction at the beginning of a movement and yet cause
also a precisely timed turn-off of the same agonist con-
traction after a given time period.
Now let us speculate on the circuit for the antago-
out the spinal cord there are reciprocal agonist/
antagonist circuits for virtually every movement that
of the basis for antagonist turn-off at the onset of
movement and then turn-on at termination of move-
bellum contains several other types of inhibitory cells
the initial inhibition of the antagonist muscles at onset
of a movement and subsequent excitation at the end
of a movement.
All these mechanisms are still partly speculation.
by which the cerebellum could cause exaggerated turn-
the degree of motor enhancement by the cerebellum
almost always incorrect for precise performance of the
tivity levels of cerebellar circuits themselves progres-
the sensitivity of the Purkinje cells to respond to the
this sensitivity change is brought about by signals from
the climbing fibers entering the cerebellum from the
inferior olivary complex.
cause extreme depolarization of the entire dendritic
strong output spike followed by a series of diminish-
joint proprioceptors will usually denote to the cere-
bellum how much the actual movement fails to match
in some way alter long-term sensitivity of the Purkinje
aspects of cerebellar control of movements approach
Motor Control
vestibulocerebellum
of the small flocculonodular cerebellar lobes (that
fractions of a second, and (2) feedback information
intended sequential plan of movement
the midbrain red nucleus, telling the cerebellum the
information when a movement is performed: (1) infor-
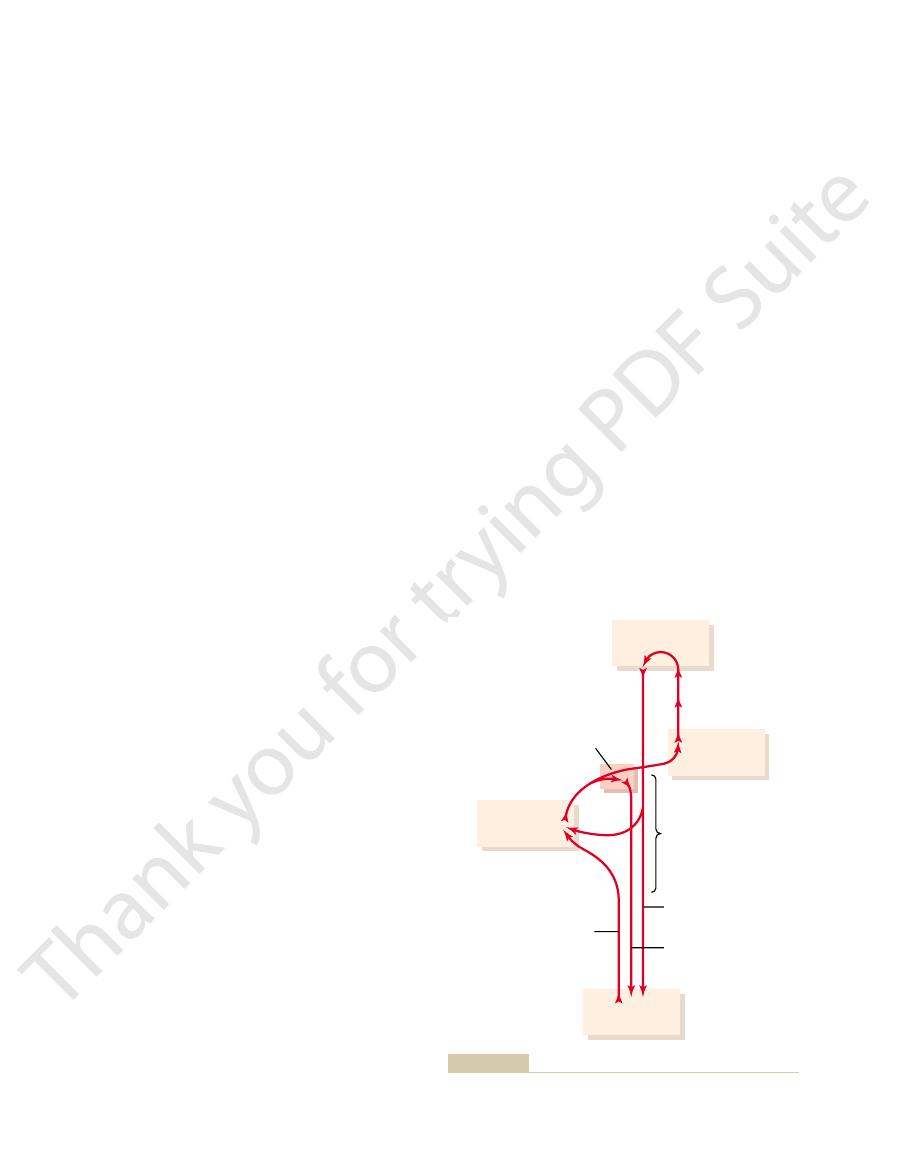
As shown in Figure 56–8, the intermediate zone of
Cerebellar Cortex and the Interposed Nucleus
Limb Movements by Way of the Intermediate
Spinocerebellum—Feedback Control of Distal
librium even during extremely rapid motion, including
Thus, during control of equilibrium, it is presumed
brain’s progression to the next sequential movement.
The results of these calculations are the key to the
different parts will be during the next few milliseconds.
directions the body parts are moving. It is then the
performed rapidly? The answer is that the signals from
sequential act, especially when the movements are
occur. How, then, is it possible for the brain to know
time. Therefore, it is never possible for return signals
15 to 20 milliseconds. The feet of a person running
to 120 m/sec in the spinocerebellar afferent tracts, the
most rapidly conducting sensory pathways are used, up
ferent parts of the body to the brain. Even when the
the vestibular apparatus.
rapid changes
muscle contractions of the spine, hips, and shoulders
late the semicircular ducts. This suggests that the
changes in direction
during stasis, especially so when these movements
cerebellar dysfunction, equilibrium is far more dis-
stem? A clue is the fact that in people with vestibulo-
We still must ask the question, what role does the
movements.
which constitute the vestibulocerebellum, causes
in Chapter 55, loss of the flocculonodular lobes and
in the inner ear developed. Furthermore, as discussed
The vestibulocerebellum originated phylogenetically
Spinal Cord to Control Equilibrium and
Vestibulocerebellum—Its Function in
development of “motor imagery” of movements
advance of the actual movements. This is called
sequential voluntary body and limb movements,
the cerebrum. It transmits its output information
to the intermediate zones. It receives virtually all
lateral zones of the cerebellar hemispheres, lateral
. This consists of the large
3. The
fingers.
portions of the limbs, especially the hands and
sides of the vermis. It provides the circuitry for
. This consists of most of the
2. The
for most of the body’s equilibrium movements.
portions of the vermis. It provides neural circuits
The Nervous System: C. Motor and Integrative Neurophysiology
704
Unit XI
lie under the posterior cerebellum) and adjacent
spinocerebellum
vermis of the posterior and anterior cerebellum
plus the adjacent intermediate zones on both
coordinating mainly movements of the distal
cerebrocerebellum
its input from the cerebral motor cortex and
adjacent premotor and somatosensory cortices of
in the upward direction back to the brain,
functioning in a feedback manner with the
cerebral cortical sensorimotor system to plan
planning these as much as tenths of a second in
to be performed.
Association with the Brain Stem and
Postural Movements
at about the same time that the vestibular apparatus
adjacent portions of the vermis of the cerebellum,
extreme disturbance of equilibrium and postural
vestibulocerebellum play in equilibrium that cannot
be provided by other neuronal machinery of the brain
turbed during performance of rapid motions than
involve
of movement and stimu-
vestibulocerebellum is especially important in con-
trolling balance between agonist and antagonist
during
in body positions as required by
One of the major problems in controlling balance is
the amount of time required to transmit position
signals and velocity of movement signals from the dif-
delay for transmission from the feet to the brain is still
rapidly can move as much as 10 inches during that
from the peripheral parts of the body to reach the
brain at the same time that the movements actually
when to stop a movement and to perform the next
the periphery tell the brain how rapidly and in which
function of the vestibulocerebellum to calculate in
advance from these rates and directions where the
that information from both the body periphery and the
vestibular apparatus is used in a typical feedback
control circuit to provide anticipatory correction of
postural motor signals necessary for maintaining equi-
rapidly changing directions of motion.
each cerebellar hemisphere receives two types of
mation from the cerebral motor cortex and from
for the next few
Motor cortex
Thalamus
Muscles
Red nucleus
Intermediate
zone of
cerebellum
Spinocerebellar
tract
Mesencephalon,
pons, and medulla
Corticospinal tract
Reticulospinal
and rubrospinal
tracts
especially the intermediate zone of the cerebellum.
Cerebral and cerebellar control of voluntary movements, involving
Figure 56–8

complex purposeful movements of the hands, fingers,
dentate nuclei, can lead to extreme incoordination of
bellar hemispheres along with their deep nuclei, the
Even so, destruction of the lateral zones of the cere-
the peripheral parts of the body. Also, almost all com-
speak. Yet the large lateral zones of the cerebellar
ment, especially with the hands and fingers, and to
enlarged. This goes along with human abilities to plan
In human beings, the lateral zones of the two cerebel-
Hemisphere to Plan, Sequence, and Time
Large Lateral Zone of the Cerebellar
Cerebrocerebellum—Function of the
for preplanned rapid ballistic movements. One also
fully organized to perform this biphasic, first excitatory
bellum as described earlier, one sees that it is beauti-
movement off. Thus, the automatism of ballistic move-
of the cerebellar circuit, the motor cortex has to think
beyond the intended mark. Therefore, in the absence
turn off, usually allowing the movement to go well
developed is weak, and (3) the movements are slow to
that the cerebellum usually provides, (2) the force
Three major changes occur: (1) The movements are
in a car.
ments of the eyes, in which the eyes jump from one
stop.Another important example is the saccadic move-
ballistic movements
over. These movements are called
fingers in typing, occur so rapidly that it is not possi-
movements of the body, such as the movements of the
control by the nervous system, the cerebellum pro-
damping circuits built into the mechanisms. For motor
This is the basic characteristic
as well as the tremor.
the intended point, thereby preventing the overshoot
But, if the cerebellum is intact, appropriate learned,
, or
on its mark. This effect is called an
intended point for several cycles before it finally fixes
tuted. Thus, the arm oscillates back and forth past its
tum, overshoots once more in the opposite direction,
intended position. But the arm, by virtue of its momen-
destroyed, the conscious centers of the cerebrum even-
. If overshooting
overshoot
ments have a tendency to
stopped. Because of momentum, all pendular move-
moved, momentum develops, and the momentum
body are “pendular.” For instance, when an arm is
ably, the olivary-Purkinje cell system along with pos-
olivary complex; if the signals do not compare favor-
dorsal spinocerebellar tract. We learned earlier that
ceptor sensory organs, transmitted principally in the
neurons, and this, too, is integrated with the signals
to the cerebellum an “efference” copy of the actual
back to the cerebellum from the periphery. In fact,
by the respective parts of the body, as transmitted
corticopontocerebellar tract, with the “performance”
higher levels of the motor control system, as transmit-
cerebellum seems to compare the “intentions” of the
forming acute purposeful patterned movements. The
vides smooth, coordinate movements of the agonist
This part of the cerebellar motor control system pro-
of the limbs, particularly the hands and fingers.
gray matter, the neurons that control the distal parts
The rubrospinal tract in turn joins
magnocellular portion
movements, the deep nuclear cells of the interposed
actual movements
the distal proprioceptors of the limbs, telling the cere-
from the peripheral parts of the body, especially from
Chapter 56
Contributions of the Cerebellum and Basal Ganglia to Overall Motor Control
705
bellum what
result.
After the intermediate zone of the cerebellum has
compared the intended movements with the actual
nucleus send corrective output signals (1) back to the
cerebral motor cortex through relay nuclei in the thal-
amus and (2) to the
(the lower
portion) of the red nucleus that gives rise to the
rubrospinal tract.
the corticospinal tract in innervating the lateral most
motor neurons in the anterior horns of the spinal cord
and antagonist muscles of the distal limbs for per-
ted to the intermediate cerebellar zone through the
the ventral spinocerebellar tract even transmits back
motor control signals that reach the anterior motor
arriving from the muscle spindles and other proprio-
similar comparator signals also go to the inferior
sibly other cerebellar learning mechanisms eventually
corrects the motions until they perform the desired
function.
Function of the Cerebellum to Prevent Overshoot of Movements
and to “Damp” Movements.
Almost all movements of the
must be overcome before the movement can be
does occur in a person whose cerebellum has been
tually recognize this and initiate a movement in the
reverse direction attempting to bring the arm to its
and appropriate corrective signals must again be insti-
action tremor
intention tremor.
subconscious signals stop the movement precisely at
of a damping system. All control systems regulating
pendular elements that have inertia must have
vides most of this damping function.
Cerebellar Control of Ballistic Movements.
Most rapid
ble to receive feedback information either from the
periphery to the cerebellum or from the cerebellum
back to the motor cortex before the movements are
,
meaning that the entire movement is preplanned and
set into motion to go a specific distance and then to
position to the next when reading or when looking at
successive points along a road as a person is moving
Much can be understood about the function of the
cerebellum by studying the changes that occur in these
ballistic movements when the cerebellum is removed.
slow to develop and do not have the extra onset surge
extra hard to turn ballistic movements on and again
has to think hard and take extra time to turn the
ments is lost.
If one considers once again the circuitry of the cere-
and then delayed inhibitory function that is required
sees that the built-in timing circuits of the cerebellar
cortex are fundamental to this particular ability of the
cerebellum.
Complex Movements
lar hemispheres are highly developed and greatly
and perform intricate sequential patterns of move-
hemispheres have no direct input of information from
munication between these lateral cerebellar areas and
the cerebral cortex is not with the primary cerebral
motor cortex itself but instead with the premotor area
and primary and association somatosensory areas.

begun; if the cerebellum is not available to do this, the
beyond the point of intention. This results from the
of the cerebellum, a person ordinarily moves the hand
Past pointing
pensatory movement.This effect is called
predict how far movements will go. Therefore, the
lum, the subconscious motor control system cannot
pointed out previously, in the absence of the cerebel-
Two of the most important symp-
, or
tion of the cerebellum, the cerebellar lesion usually
Therefore, to cause serious and continuing dysfunc-
. Thus, the remaining portions of the motor
slowly
as long as the animal performs all movements
bellar nuclei are not removed along with the cortex, the
side of the brain has been removed, if the deep cere-
ties in motor function. In fact, several months after as
Clinical Abnormalities of
rapidly changing
can be compared; it is often stated that the cerebellum
base,” perhaps using time-delay circuits, against which
is quite possible that the cerebellum provides a “time-
extramotor predictive functions of the cerebellum. It
We are only now beginning to learn about these
tions of the cerebellum in monkeys. Such a monkey
approaching an object. A striking experiment that
pation. As an example, a person can predict from
the brain, but both of these require cerebellar partici-
body. For instance, the rates of progression of both
helps to “time” events other than movements of the
The cerebrocerebellum (the large lateral lobes) also
smooth progression of movements
required for writing, running, or even talking) to
too late. Therefore, lesions in the lateral zones of the
ceeding movement may begin too early or, more likely,
tial movement needs to begin. As a result, the suc-
given time. Without this timing capability, the person
ment. In the absence of these cerebellar zones, one
Timing Function.
disturbed for rapid movements.
the cerebellar hemispheres, this capability is seriously
succession. In the absence of the large lateral zones of
of normal motor function is one’s ability to progress
To summarize, one of the most important features
seconds later.
movement
present movement is still occurring. Thus, the lateral
between cerebellum and cerebral cortex, appropriate
lar hemispheres. Then, amid much two-way traffic
tor areas of the cerebral cortex, and from there the
basal ganglia. It seems that the “plan” of sequential
The planning of
movements.
tial movements and (2) the “timing” of the sequential
aspects of motor control: (1) the planning of sequen-
the primary motor cortex. However, experimental
and feet and of the speech apparatus. This has been
The Nervous System: C. Motor and Integrative Neurophysiology
706
Unit XI
difficult to understand because of lack of direct com-
munication between this part of the cerebellum and
studies suggest that these portions of the cerebellum
are concerned with two other important but indirect
Planning of Sequential Movements.
sequential movements requires that the lateral zones
of the hemispheres communicate with both the pre-
motor and the sensory portions of the cerebral cortex,
and it requires two-way communication between these
cerebral cortex areas with corresponding areas of the
movements actually begins in the sensory and premo-
plan is transmitted to the lateral zones of the cerebel-
motor signals provide transition from one sequence of
movements to the next.
An exceedingly interesting observation that sup-
ports this view is that many neurons in the cerebellar
dentate nuclei display the activity pattern for the
sequential movement that is yet to come while the
cerebellar zones appear to be involved not with what
movement is happening at a given moment but with
what will be happening during the next sequential
a fraction of a second or perhaps even
smoothly from one movement to the next in orderly
Another important function of the
lateral zones of the cerebellar hemispheres is to
provide appropriate timing for each succeeding move-
loses the subconscious ability to predict ahead of time
how far the different parts of the body will move in a
becomes unable to determine when the next sequen-
cerebellum cause complex movements (such as those
become incoordinate and lacking ability to progress in
orderly sequence from one movement to the next.
Such cerebellar lesions are said to cause failure of
.
Extramotor Predictive Functions of the Cerebrocerebellum.
auditory and visual phenomena can be predicted by
the changing visual scene how rapidly he or she is
demonstrates the importance of the cerebellum in this
ability is the effects of removing the large lateral por-
occasionally charges the wall of a corridor and literally
bashes its brains because it is unable to predict when
it will reach the wall.
signals from other parts of the central nervous system
is particularly helpful in interpreting
spatiotemporal relations in sensory information.
the Cerebellum
An important feature of clinical cerebellar abnormali-
ties is that destruction of small portions of the lateral
cerebellar cortex seldom causes detectable abnormali-
much as one half of the lateral cerebellar cortex on one
motor functions of the animal appear to be almost
normal
control system are capable of compensating tremen-
dously for loss of parts of the cerebellum.
must involve one or more of the deep cerebellar
nuclei—the dentate, interposed
fastigial nuclei.
Dysmetria and Ataxia.
toms of cerebellar disease are dysmetria and ataxia. As
movements ordinarily overshoot their intended mark;
then the conscious portion of the brain overcompen-
sates in the opposite direction for the succeeding com-
dysmetria, and
it results in uncoordinated movements that are called
ataxia. Dysmetria and ataxia can also result from lesions
in the spinocerebellar tracts because feedback informa-
tion from the moving parts of the body to the cerebel-
lum is essential for cerebellar timing of movement
termination.
Past Pointing.
means that in the absence
or some other moving part of the body considerably
fact that normally the cerebellum initiates most of the
motor signal that turns off a movement after it is
movement ordinarily goes beyond the intended mark.

hemispheres. Note also that almost all motor and
lateral to and surrounding the thalamus, occupying a
. They are located mainly
side of the brain, these ganglia consist of the
basal ganglia to other structures of the brain. On each
Figure 56–9 shows the anatomical relations of the
cortex and corticospinal motor control system. In fact,
The basal ganglia, like the cerebellum, constitute
the side of the cerebellar lesion. The hypotonia results
larly of the dentate and interposed nuclei, causes
Loss of the deep cerebellar nuclei, particu-
lar cerebellum from the semicircular ducts.
lar lobes of the cerebellum are damaged; in this instance
cerebellum. It occurs especially when the flocculonodu-
movements of the eyes rather than steady fixation, and
off-center type of fixation results in rapid, tremulous
fixate the eyes on a scene to one side of the head. This
motor movements.
ing and failure of the cerebellar system to “damp” the
action tremor,
mark. This reaction is called an
mark, first overshooting the mark and then vibrating
to oscillate, especially when they approach the intended
bellum performs a voluntary act, the movements tend
When a person who has lost the cere-
Intention Tremor.
ble. This is called
intervals, and resultant speech that is often unintelligi-
weak, some held for long intervals, some held for short
jumbled vocalization, with some syllables loud, some
piratory system. Lack of coordination among these and
vidual muscle movements in the larynx, mouth, and res-
dysdiadochokine-
and downward motions. This is called
result, a series of stalled attempted but jumbled move-
the hand during any portion of the movement. As a
“loses” all perception of the instantaneous position of
and downward at a rapid rate. The patient rapidly
can occur. One can demonstrate this readily by having
too late, so that no orderly “progression of movement”
during rapid motor movements. As a result, the suc-
be at a given time, it “loses” perception of the parts
When the motor control system
Therefore, past pointing is actually a manifestation of
Chapter 56
Contributions of the Cerebellum and Basal Ganglia to Overall Motor Control
707
dysmetria.
Failure of Progression
Dysdiadochokinesia.
fails to predict where the different parts of the body will
ceeding movement may begin much too early or much
a patient with cerebellar damage turn one hand upward
ments occurs instead of the normal coordinate upward
sia.
Dysarthria.
Another example in which failure of pro-
gression occurs is in talking because the formation of
words depends on rapid and orderly succession of indi-
inability to adjust in advance either the intensity of
sound or duration of each successive sound causes
dysarthria.
back and forth several times before settling on the
intention tremor or an
and it results from cerebellar overshoot-
Cerebellar Nystagmus.
Cerebellar nystagmus is tremor of
the eyeballs that occurs usually when one attempts to
it is another manifestation of failure of damping by the
it is also associated with loss of equilibrium because of
dysfunction of the pathways through the flocculonodu-
Hypotonia.
decreased tone of the peripheral body musculature on
from loss of cerebellar facilitation of the motor cortex
and brain stem motor nuclei by tonic signals from the
deep cerebellar nuclei.
Basal Ganglia—Their Motor
Functions
another accessory motor system that functions usually
not by itself but in close association with the cerebral
the basal ganglia receive most of their input signals
from the cerebral cortex itself and also return almost
all their output signals back to the cortex.
caudate
nucleus, putamen, globus pallidus, substantia nigra,
and subthalamic nucleus
large portion of the interior regions of both cerebral
Longitudinal fissure
Caudate nucleus
Tail of caudate
LATERAL
Fibers to and from
spinal cord in
internal capsule
Putamen and
globus pallidus
POSTERIOR
ANTERIOR
Thalamus
and Physiology. Philadelphia: WB
AC: Basic Neuroscience: Anatomy
sional view. (Redrawn from Guyton
thalamus, shown in three-dimen-
ganglia to the cerebral cortex and
Anatomical relations of the basal
Figure 56–9
Saunders Co, 1992.)
primary motor cortex. Thus,
the thalamus, and finally return to the cerebral primary
to the internal portion of the globus pallidus, next to
putamen (mainly bypassing the caudate nucleus), then
areas of the sensory cortex. Next they pass to the
They begin mainly in the premotor and supplementary
Figure 56–11
most of them performed subconsciously.
eyes, and virtually any other of our skilled movements,
aspects of vocalization, controlled movements of the
ing a baseball, the movements of shoveling dirt, most
a basketball through a hoop, passing a football, throw-
cutting paper with scissors, hammering nails, shooting
time how to write.
becomes crude, as if one were learning for the first
longer provide these patterns. Instead, one’s writing
ganglia, the cortical system of motor control can no
alphabet. When there is serious damage to the basal
. An example is the writing of letters of the
Executing Patterns of Motor Activity—
Function of the Basal Ganglia in
cially on two major circuits, the
basal ganglia.
cuitry of the basal ganglia system, showing the tremen-
and cerebellar circuitry. To the right is the major cir-
the motor cortex, thalamus, and associated brain stem
complex, as shown in Figure 56–10. To the left is shown
The anatomical
Neuronal Circuitry of the Basal Ganglia.
of the brain. It is important for our
. This space is called
between the major masses of the basal ganglia, the
The Nervous System: C. Motor and Integrative Neurophysiology
708
Unit XI
sensory nerve fibers connecting the cerebral cortex
and spinal cord pass through the space that lies
caudate nucleus and the putamen
the internal capsule
current discussion because of the intimate association
between the basal ganglia and the corticospinal system
for motor control.
connections between the basal ganglia and the other
brain elements that provide motor control are
dous interconnections among the basal ganglia
themselves plus extensive input and output pathways
between the other motor regions of the brain and the
In the next few sections we will concentrate espe-
putamen circuit and the
caudate circuit.
The Putamen Circuit
One of the principal roles of the basal ganglia in motor
control is to function in association with the corti-
cospinal system to control complex patterns of motor
activity
Other patterns that require the basal ganglia are
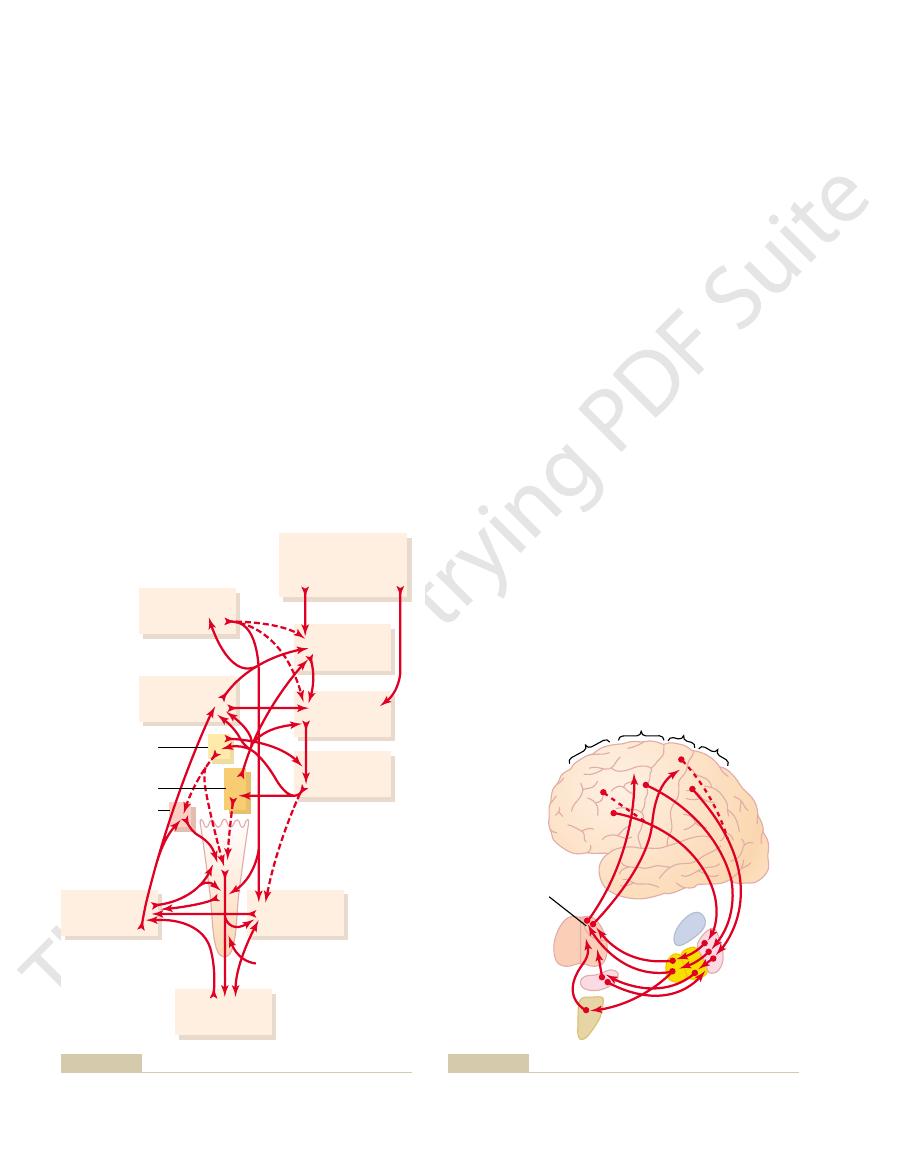
Neural Pathways of the Putamen Circuit.
shows the principal pathways through the basal
ganglia for executing learned patterns of movement.
areas of the motor cortex and in the somatosensory
the ventroanterior and ventrolateral relay nuclei of
motor cortex and to portions of the premotor and sup-
plementary cerebral areas closely associated with the
the putamen circuit has its
Motor cortex
Thalamus
Muscles
Globus
pallidus
Inferior olive
Reticular formation
Cerebellum
Putamen
Caudate
nucleus
Premotor and
supplemental motor
association areas
Subthalamus
Substantia nigra
Red nucleus
cerebellar system for movement control.
Relation of the basal ganglial circuitry to the corticospinal-
Figure 56–10
Caudate
Subthalamus
Substantia nigra
Premotor and
supplemental Primary motor
Prefrontal
Ventroanterior and
ventrolateral
nuclei of thalamus
Putamen
Globus
pallidus
internal/external
Somatosensory
cution of learned patterns of movement.
Putamen circuit through the basal ganglia for subconscious exe-
Figure 56–11

write a small “a” on a piece of paper or a large “a” on
the letter “a” slowly or rapidly. Also, he or she may
the movement will be. For instance, a person may write
Two important capabilities of the brain in controlling
Intensity of Movements
Change the Timing and to Scale the
Function of the Basal Ganglia to
a complex goal that might itself last for many seconds.
determines subconsciously, and within seconds, which
appropriately. Thus, cognitive control of motor activity
thinking for too long a time, to respond quickly and
might not have the instinctive knowledge, without
a tree. Without the cognitive functions, the person
(2) beginning to run, and (3) even attempting to climb
ual muscle movements.
primary motor cortex. Instead, the returning signals go
motor areas of the cerebral cortex, but with almost
back to the prefrontal, premotor, and supplementary
ventroanterior and ventrolateral thalamus, and finally
internal globus pallidus, then to the relay nuclei of the
the caudate nucleus, they are next transmitted to the
patterns.
of the cerebral cortex overlying the caudate nucleus,
poral lobes. Furthermore, the caudate nucleus receives
curving forward again like the letter “C” into the tem-
through the parietal and occipital lobes, and finally
riorly in the frontal lobes, then passing posteriorly
extends into all lobes of the cerebrum, beginning ante-
is that the caudate nucleus, as shown in Figure 56–9,
those of the putamen circuit. Part of the reason for this
shown in Figure 56–12, are somewhat different from
The neural connections between the caudate
role in this cognitive control of motor activity.
. The caudate nucleus plays a major
erated in the mind, a process called
information already stored in memory. Most of our
the brain, using both sensory input to the brain plus
The term
Cognitive Control of Sequences of
Role of the Basal Ganglia for
discuss in more detail later.
, which we
Parkinson
, and
chorea
the body, called
in the hands, face, and other parts of
ing movements
flick-
of an entire limb, a condition called
flailing movements
athetosis.
a hand, an arm, the neck, or the face—movements
writhing movements
movement become severely abnormal. For instance,
the circuit is damaged or blocked, certain patterns of
answer is poorly known. However, when a portion of
tion to help execute patterns of movement? The
to the motor cortex by way of the thalamus.
thalamus, and the substantia nigra—finally returning
putamen through the external globus pallidus, the sub-
cortex. Func-
primary motor cortex itself. Then its outputs do go
Chapter 56
Contributions of the Cerebellum and Basal Ganglia to Overall Motor Control
709
inputs mainly from those parts of the brain adjacent to
the primary motor cortex but not much from the
mainly back to the primary motor cortex or closely
associated premotor and supplementary
tioning in close association with this primary putamen
circuit are ancillary circuits that pass from the
Abnormal Function in the Putamen Circuit: Athetosis, Hemibal-
lismus, and Chorea.
How does the putamen circuit func-
lesions in the globus pallidus frequently lead to spon-
taneous and often continuous
of
called
A lesion in the subthalamus often leads to sudden
hemiballismus.
Multiple small lesions in the putamen lead to
.
Lesions of the substantia nigra lead to the common
and extremely severe disease of rigidity, akinesia
tremors known as
’s disease
Motor Patterns—The Caudate Circuit
cognition means the thinking processes of
motor actions occur as a consequence of thoughts gen-
cognitive control
of motor activity
nucleus and the corticospinal motor control system,
large amounts of its input from the association areas
mainly areas that also integrate the different types of
sensory and motor information into usable thought
After the signals pass from the cerebral cortex to
none of the returning signals passing directly to the
to those accessory motor regions in the premotor and
supplementary motor areas that are concerned with
putting together sequential patterns of movement
lasting 5 or more seconds instead of exciting individ-
A good example of this would be a person seeing a
lion approach and then responding instantaneously
and automatically by (1) turning away from the lion,
patterns of movement will be used together to achieve
movement are (1) to determine how rapidly the move-
ment is to be performed and (2) to control how large
Caudate
Subthalamus
Substantia nigra
Premotor and
supplemental Primary motor
Prefrontal
Ventroanterior and
ventrolateral
nuclei of thalamus
Putamen
Globus
pallidus
internal/external
Somatosensory
of sequential and parallel motor patterns to achieve specific con-
Caudate circuit through the basal ganglia for cognitive planning
Figure 56–12
scious goals.

inhibitory agent. Therefore, GABA neurons in the
the neurotransmitter GABA always functions as an
For the present, it should be remembered that
we discuss behavior, sleep, wakefulness, and functions
basal ganglia, as well as in subsequent chapters when
inhibitory transmitters. We will have more to say about
especially by the dopamine, GABA, and serotonin
tamate pathways
the cerebrum. In addition to all these are
, and several other neurotrans-
nucleus and putamen, and (4) multiple general path-
choline
the globus pallidus and substantia nigra, (3)
gamma-aminobutyric acid (GABA)
and putamen, (2)
within the basal ganglia, showing (1)
Figure 56–14 demonstrates the interplay of several
Functions of Specific
circuit. However, our understanding of function in the
her right body for the performance of tasks, almost not
or her right arm, right hand, or other portions of his or
Also, such a person will try always to avoid using his
another human being, providing proper proportions
Figure 56–13 shows the way in which a person lacking
tion of the body and its parts to all its surroundings.
association with the cerebral cortex. One especially
ganglia do not function alone; they function in close
sometimes they are nonexistent. Here again, the basal
these timing and scaling functions are poor; in fact,
same.
a chalkboard. Regardless of the choice, the propor-
The Nervous System: C. Motor and Integrative Neurophysiology
710
Unit XI
tional characteristics of the letter remain nearly the
In patients with severe lesions of the basal ganglia,
important cortical area is the posterior parietal cortex,
which is the locus of the spatial coordinates for motor
control of all parts of the body as well as for the rela-
a left posterior parietal cortex might draw the face of
for the right side of the face but almost ignoring the
left side (which is in his or her right field of vision).
knowing that these parts of his or her body exist.
Because the caudate circuit of the basal ganglial
system functions mainly with association areas of the
cerebral cortex such as the posterior parietal cortex,
presumably the timing and scaling of movements are
functions of this caudate cognitive motor control
basal ganglia is still so imprecise that much of what is
conjectured in the last few sections is analytical deduc-
tion rather than proven fact.
Neurotransmitter Substances in the
Basal Ganglial System
specific neurotransmitters that are known to function
dopamine path-
ways from the substantia nigra to the caudate nucleus
pathways from the caudate nucleus and putamen to
acetyl-
pathways from the cortex to the caudate
ways from the brain stem that secrete norepinephrine,
serotonin, enkephalin
mitters in the basal ganglia as well as in other parts of
multiple glu-
that provide most of the excitatory
signals (not shown in the figure) that balance out
the large numbers of inhibitory signals transmitted
some of these neurotransmitter and hormonal systems
in subsequent sections when we discuss diseases of the
of the autonomic nervous system.
feedback loops from the cortex through the basal
nates of the right field of vision are stored.
damage in his or her left parietal cortex where the spatial coordi-
Typical drawing that might be made by a person who has severe
Figure 56–13
Caudate nucleus
From cortex
From brain stem
Substantia
nigra
1. Norepinephrine
2. Serotonin
3. Enkephalin
Putamen
Globus
pallidus
Dopamine
GABA
Ach
Neuronal pathways that secrete different types of neurotransmit-
Figure 56–14
ter substances in the basal ganglia. Ach, acetylcholine; GABA,
gamma-aminobutyric acid.

loss of acetylcholine-secreting neurons, perhaps espe-
not result from the loss of GABA neurons but from the
The dementia in Huntington’s disease probably does
the distortional movements.
globus pallidus and substantia nigra. This loss of inhibi-
of the GABA neurons normally inhibit portions of the
neurons in many parts of the brain. The axon terminals
bodies of the GABA-secreting neurons in the caudate
The abnormal movements of Huntington’s disease
dementia develops along with the motor dysfunctions.
movements of the entire body. In addition, severe
usually begins causing symptoms at age 30 to 40 years.
Huntington’s disease is a hereditary disorder that
Huntington’s Disease (Huntington’s Chorea)
results.
amus have been used, sometimes with surprisingly good
with Parkinson’s disease, lesions placed in the subthal-
sometimes serious neurological damage. In monkeys
rior nuclei of the thalamus, which blocked part of the
these signals surgically. For a number of years, surgical
abnormalities in Parkinson’s disease, multiple attempts
in the Basal Ganglia.
Treatment by Destroying Part of the Feedback Circuitry
would become the treatment of the future.
months. If persistence could be achieved, perhaps this
However, the cells do not live for more than a few
short-term success to treat Parkinson’s disease.
Transplantation of dopamine-secreting cells (cells
Treatment with Transplanted Fetal Dopamine Cells.
of one of these drugs alone.
neurons in the substantia nigra. Therefore, appropriate
In addition, for reasons not understood, this treatment
remains in the basal ganglial tissues for a longer time.
secreted. Therefore, any dopamine that is released
inhibits monoamine oxidase, which is responsible for
-deprenyl. This drug
Parkinson’s disease is the drug
Treatment with l-Deprenyl.
-dopa does allow it to pass.
brain barrier, even though the slightly different struc-
putamen. Administration of dopamine itself does not
-dopa is converted in the brain into dopamine, and the
and akinesia. The reason for this is believed to be that
liorates many of the symptoms, especially the rigidity
to patients with Parkinson’s disease usually ame-
Treatment with l-Dopa.
drive for motor activity so greatly that akinesia results.
decreased along with its decrease in the basal ganglia. It
, is often
system, especially in the
ulative. However, dopamine secretion in the limbic
instead of smooth. The cause of this akinesia is still spec-
occur, they are usually stiff and staccato in character
the patient’s willpower. Then, when the movements do
The mental effort, even mental anguish, that is necessary
even the simplest movement in severe parkinsonism, the
toms of muscle rigidity and tremor, because to perform
that occurs in Parkinson’s disease is often
The
intention tremor.
bellar tremor, which occurs only when the person per-
, in contradistinction to cere-
of Parkinson’s disease. This
bition, leading to the
rigidity.
many or all of the muscles of the body, thus leading to
control system. These signals could overly excite
ter; therefore, destruction of the dopaminergic neurons
unknown. However, the dopamine secreted in the
The causes of these abnormal motor effects are
second, and (3) serious difficulty in initiating movement,
person is resting at a fixed rate of 3 to 6 cycles per
ity of much of the musculature of the body, (2) invol-
and putamen. The disease is characterized by (1) rigid-
Parkinson’s disease, known also as
Parkinson’s Disease
Parkinson’s disease and Huntington’s disease.
eases result from damage in the basal ganglia. These are
globus pallidus and subthalamus, two other major dis-
, which have
under some conditions.
so that it, too, undoubtedly functions as a stabilizer
motor control systems. Dopamine also functions as an
positive feedback loops, thus lending stability to the
, rather than
negative feedback loops
Chapter 56
Contributions of the Cerebellum and Basal Ganglia to Overall Motor Control
711
ganglia and then back to the cortex make virtually
all these loops
inhibitory neurotransmitter in most parts of the brain,
Clinical Syndromes Resulting from
Damage to the Basal Ganglia
Aside from athetosis and hemiballismus
already been mentioned in relation to lesions in the
paralysis agitans,
results from widespread destruction of that portion of
the substantia nigra (the pars compacta) that sends
dopamine-secreting nerve fibers to the caudate nucleus
untary tremor of the involved areas even when the
called akinesia.
caudate nucleus and putamen is an inhibitory transmit-
in the substantia nigra of the parkinsonian patient the-
oretically would allow the caudate nucleus and putamen
to become overly active and possibly cause continuous
output of excitatory signals to the corticospinal motor
Some of the feedback circuits might easily oscillate
because of high feedback gains after loss of their inhi-
tremor
tremor is quite different from that of cerebellar disease
because it occurs during all waking hours and therefore
is an involuntary tremor
forms intentionally initiated movements and therefore
is called
akinesia
much more distressing to the patient than are the symp-
person must exert the highest degree of concentration.
to make the desired movements is often at the limit of
nucleus accumbens
has been suggested that this might reduce the psychic
Administration of the drug
L
-
dopa
L
dopamine then restores the normal balance between
inhibition and excitation in the caudate nucleus and
have the same effect because dopamine has a chemical
structure that will not allow it to pass through the blood-
ture of
L
Another treatment for
L
destruction of most of the dopamine after it has been
helps to slow destruction of the dopamine-secreting
combinations of
L
-dopa therapy along with
L
-deprenyl
therapy usually provide much better treatment than use
obtained from the brains of aborted fetuses) into the
caudate nuclei and putamen has been used with some
Because abnormal signals from
the basal ganglia to the motor cortex cause most of the
have been made to treat these patients by blocking
lesions were made in the ventrolateral and ventroante-
feedback circuit from the basal ganglia to the cortex;
variable degrees of success were achieved—as well as
It is characterized at first by flicking movements in indi-
vidual muscles and then progressive severe distortional
are believed to be caused by loss of most of the cell
nucleus and putamen and of acetylcholine-secreting
tion is believed to allow spontaneous outbursts of
globus pallidus and substantia nigra activity that cause
cially in the thinking areas of the cerebral cortex.

situation, such as planning one’s immediate motor
cerebral and basal ganglia circuit, beginning in the
thus controlling dimensions of the patterns.
these patterns for writing small or writing very large,
typing. Also, the basal ganglia are required to modify
letters of the alphabet, for throwing a ball, and for
The types of motor patterns that require the basal
learned patterns of movement
different from those of the cerebellum. Their most
The basal
slow and calculated movements can still occur, but it
movements have to be rapid. Without the cerebellum,
The cerebellum functions mainly when muscle
The neural circuit for this passes from the cerebral
direction, all this occurring in a fraction of a second.
still another level of motor control: it helps to program
The cerebellum functions with the cerebral cortex at
learned with experience.
movement at the intended point. Furthermore, there
ment, the
at the start of a movement. Near the end of each move-
accessory motor functions, especially to provide extra
At the cerebral cortex level, the cerebellum oper-
mal oscillations.
At the brain stem level, the cerebellum functions to
an unexpectedly heavy load, a long stretch reflex signal
reflex, so that when a contracting muscle encounters
functions with all levels of muscle control. It functions
The cerebellum
wired.”
determined by heredity and are said to be “hard
can be “learned,” whereas cord patterns are mainly
The cortical patterns usually are complex; also, they
bypass the cord patterns, replacing them with higher-
teristics. When needed, the corticospinal system can
action. It can also change the intensities of the differ-
vating motor signals to the spinal cord. It functions
The motor cortex system provides most of the acti-
Motor Cortex Level
general motor control of the body: (1) maintenance of
The hindbrain provides two major functions for
into action by higher levels of motor control, or they
in four-legged animals.
for walking, plus reciprocal motions on opposite sides
any part of the body away from a source of pain. The
instance, programmed withdrawal reflexes that pull
known about overall control of movement. To do this,
Finally, we need to summarize as best we can what is
Control System
Parts of the Total Motor
toms. How this protein causes the disease effects is now
CAG, that codes for multiple extra
has been found; it has a many-times-repeating codon,
The abnormal gene that causes Huntington’s disease
The Nervous System: C. Motor and Integrative Neurophysiology
712
Unit XI
glutamine amino
acids in the molecular structure of an abnormal neu-
ronal cell protein called huntingtin that causes the symp-
the question for major research effort.
Integration of the Many
let us first give a synopsis of the different levels of
control.
Spinal Level
Programmed in the spinal cord are local patterns of
movement for all muscle areas of the body—for
cord is the locus also of complex patterns of rhythmi-
cal motions such as to-and-fro movement of the limbs
of the body or of the hindlimbs versus the forelimbs
All these programs of the cord can be commanded
can be inhibited while the higher levels take over
control.
Hindbrain Level
axial tone of the body for the purpose of standing and
(2) continuous modification of the degrees of tone in
the different muscles in response to information from
the vestibular apparatuses for the purpose of main-
taining body equilibrium.
partly by issuing sequential and parallel commands
that set into motion various cord patterns of motor
ent patterns or modify their timing or other charac-
level patterns from the brain stem or cerebral cortex.
Associated Functions of the Cerebellum.
with the spinal cord especially to enhance the stretch
transmitted all the way through the cerebellum and
back again to the cord strongly enhances the load-
resisting effect of the basic stretch reflex.
make the postural movements of the body—especially
the rapid movements required by the equilibrium
system—smooth and continuous and without abnor-
ates in association with the cortex to provide many
motor force for turning on muscle contraction rapidly
cerebellum turns on antagonist muscles at
exactly the right time and with proper force to stop the
is good physiologic evidence that all aspects of this
turn-on/turn-off patterning by the cerebellum can be
in advance muscle contractions that are required for
smooth progression from a present rapid movement in
one direction to the next rapid movement in another
cortex to the large lateral zones of the cerebellar hemi-
spheres and then back to the cerebral cortex.
is difficult for the corticospinal system to achieve rapid
and changing intended movements to execute a par-
ticular goal or especially to progress smoothly from
one rapid movement to the next.
Associated Functions of the Basal Ganglia.
ganglia are essential to motor control in ways entirely
important functions are (1) to help the cortex execute
subconscious but
and (2)
to help plan multiple parallel and sequential patterns
of movement that the mind must put together to
accomplish a purposeful task.
ganglia include those for writing all the different
At a still higher level of control is another combined
thinking processes of the cerebrum to provide overall
sequential steps of action for responding to each new
response to an assailant who hits the person in the face

Sethi KD: Tremor. Curr Opin Neurol 16:481, 2003.
Brain Res Brain Res Rev 44:103, 2004.
neuromodulation: towards a functional understanding.
Schweighofer N, Doya K, Kuroda S: Cerebellar aminergic
363:1783, 2004.
Samii A, Nutt JG, Ransom BR: Parkinson’s disease. Lancet
Parkinson’s disease. Annu Rev Med 55:41, 2004.
Olanow CW: The scientific basis for the current treatment of
N Y Acad Sci 978:425, 2002.
understand the cerebellum well enough to build one. Ann
Ohyama T, Medina JF, Nores WL, Mauk MD: Trying to
Neurology 62(1 Suppl 1):S17, 2004.
of dopaminergic innervation and basal ganglia circuits.
of motor fluctuations in Parkinson’s disease: importance
Obeso JA, Rodriguez-Oroz M, Marin C, et al: The origin
topathy? Trends Mol Med 9:414, 2003.
Li JY, Plomann M, Brundin P: Huntington’s disease: a synap-
lorubrospinal system. Physiol Rev 74:509, 1994.
Keifer J, Houk JC: Motor function of the cerebel-
Science, 4th ed. New York: McGraw-Hill, 2000.
Kandel ER, Schwartz JH, Jessell TM: Principles of Neural
81:1143, 2001.
signal transduction, and functional roles. Physiol Rev
Ito M: Cerebellar long-term depression: characterization,
perception and action. Curr Opin Neurobiol 14:203,
vision: separate but interacting cortical pathways for
Goodale MA, Westwood DA: An evolving view of duplex
discharge. Ann N Y Acad Sci 978:219, 2002.
Gibson AR, Horn KM, Pong M: Inhibitory control of olivary
of cerebellar neuronal circuitry. News Physiol Sci 13:26,
Garwicz M, Ekerot C-F, Jorntell H: Organizational principles
Elble RJ: Origins of tremor. Lancet 355:1113, 2000.
ture to Control. Amsterdam: Elsevier, 1997.
de Zeeuw CI, Strata P,Voogd J:The Cerebellum: From Struc-
302:830, 2003.
early detection of neurodegenerative disorders. Science
DeKosky ST, Marek K: Looking backward to move forward:
Bethesda: American Physiological Society, 1981, p 877.
movement. In: Handbook of Physiology, Sec. 1, Vol. II.
Brooks VB, Thach WT: Cerebellar control of posture and
eases and oxidative stress. Nat Rev Drug Discov 3:205,
Barnham KJ, Masters CL, Bush AI: Neurodegenerative dis-
update. Curr Opin Neurol 16:507, 2003.
Albin RL: Dominant ataxias and Friedreich ataxia: an
of the brain. We discuss this system in detail in
These areas are collectively called the
anterior to the hypothalamus and thalamus, and even
hypothalamus, amygdala, hippocampus, septal region
anterior, and lateral to the thalamus—including the
Basically, the brain has an older core located beneath,
play our trains of movement? We are beginning to
What is it that arouses us from inactivity and sets into
What Drives Us to Action?
embrace.
or one’s sequential response to an unexpectedly fond
Chapter 56
Contributions of the Cerebellum and Basal Ganglia to Overall Motor Control
713
learn about the motivational systems of the brain.
old regions of the thalamus and cerebral cortex them-
selves—all of which function together to initiate most
motor and other functional activities of the brain.
limbic system
Chapter 58.
References
2004.
1998.
2004.